The primary visual pathway consists of a relay system, beginning at the retina, whose ganglion cell axons form the optic nerve. The optic nerve fibers from each eye hemidecussate in the optic chiasm (OC), with nasal fibers joining the temporal fibers of the contralateral nerve. The nasal fibers continue as the optic tract on each side, synapsing with the lateral geniculate nucleus (LGN) of the thalamus. Signals are then transmitted to the primary visual cortex of the occipital lobe. The right and left visual fields are processed by opposite hemispheres. Lesions along the pathway result in vision loss or visual field deficits. Based on the type of presentation, the location of the lesion in the pathway can be ascertained.
Last updated: Dec 15, 2025
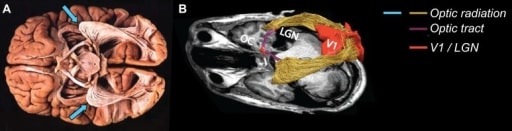
A: postmortem dissection of the optic radiation, showing the Meyer’s loop (blue arrows) extending anteriorly before turning posteriorly, going to the primary visual cortex (adapted from Sherbondy et al., 2008b); B: in vivo dissection of the optic radiation showing the OC, the optic tract (purple), LGN, Meyer’s loop (yellow), and the primary visual cortex (V1; red) (Ogawa et al., 2014)
Image: “The visual white matter: The application of diffusion MRI and fiber tractography to vision science ” by the Journal of Vision. License: CC BY 4.0.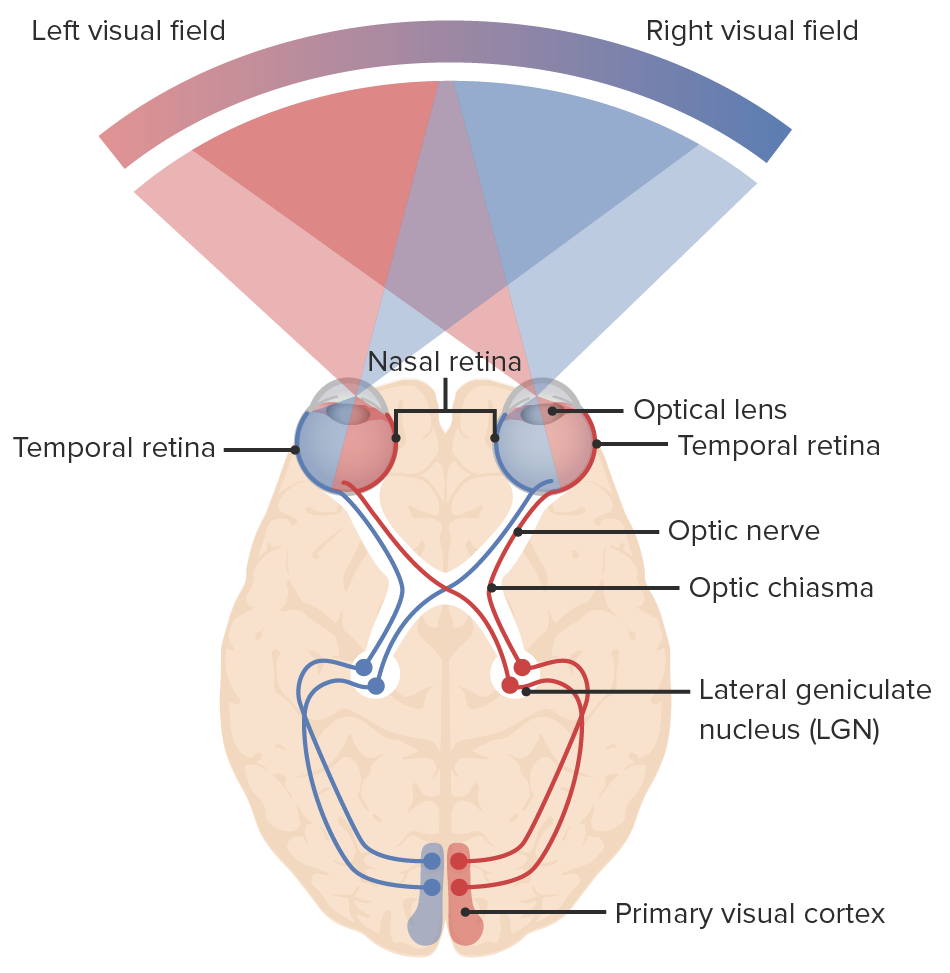
Diagram of the visual pathway and the visual fields: light enters the eye, sending signals to the retina and through the optic nerve. The nasal fibers of each eye decussate at the optic chiasm, continuing to the optic tract with the temporal fibers: right nasal fibers join the left temporal fibers (blue lines) and the left nasal fibers join the right temporal fibers (red lines). Neurons synapse at the lateral geniculate nucleus. Optic radiations connect the lateral geniculate nucleus to the primary visual cortex of the occipital lobe where visual information is processed.
Image by Lecturio.| Site of lesion | Visual field defect | Description | Possible cause |
|---|---|---|---|
| Macula Macula An oval area in the retina, 3 to 5 mm in diameter, usually located temporal to the posterior pole of the eye and slightly below the level of the optic disk. It is characterized by the presence of a yellow pigment diffusely permeating the inner layers, contains the fovea centralis in its center, and provides the best phototropic visual acuity. It is devoid of retinal blood vessels, except in its periphery, and receives nourishment from the choriocapillaris of the choroid. Eye: Anatomy (central retina Retina The ten-layered nervous tissue membrane of the eye. It is continuous with the optic nerve and receives images of external objects and transmits visual impulses to the brain. Its outer surface is in contact with the choroid and the inner surface with the vitreous body. The outermost layer is pigmented, whereas the inner nine layers are transparent. Eye: Anatomy) | Central scotoma Scotoma A localized defect in the visual field bordered by an area of normal vision. This occurs with a variety of eye diseases (e.g., retinal diseases and glaucoma); optic nerve diseases, and other conditions. Cranial Nerve Palsies | Ipsilateral central vision loss Central Vision Loss Macular Degeneration | Optic neuritis Optic neuritis Inflammation of the optic nerve. Commonly associated conditions include autoimmune disorders such as multiple sclerosis, infections, and granulomatous diseases. Clinical features include retro-orbital pain that is aggravated by eye movement, loss of color vision, and contrast sensitivity that may progress to severe visual loss, an afferent pupillary defect (Marcus-Gunn pupil), and in some instances optic disc hyperemia and swelling. Inflammation may occur in the portion of the nerve within the globe (neuropapillitis or anterior optic neuritis) or the portion behind the globe (retrobulbar neuritis or posterior optic neuritis). Cranial Nerve Palsies, retrobulbar neuritis Retrobulbar neuritis Inflammation of the optic nerve. Commonly associated conditions include autoimmune disorders such as multiple sclerosis, infections, and granulomatous diseases. Clinical features include retro-orbital pain that is aggravated by eye movement, loss of color vision, and contrast sensitivity that may progress to severe visual loss, an afferent pupillary defect (Marcus-Gunn pupil), and in some instances optic disc hyperemia and swelling. Inflammation may occur in the portion of the nerve within the globe (neuropapillitis or anterior optic neuritis) or the portion behind the globe (retrobulbar neuritis or posterior optic neuritis). Cranial Nerve Palsies |
| Optic nerve Optic nerve The 2nd cranial nerve which conveys visual information from the retina to the brain. The nerve carries the axons of the retinal ganglion cells which sort at the optic chiasm and continue via the optic tracts to the brain. The largest projection is to the lateral geniculate nuclei; other targets include the superior colliculi and the suprachiasmatic nuclei. Though known as the second cranial nerve, it is considered part of the central nervous system. The 12 Cranial Nerves: Overview and Functions | Ipsilateral monocular blindness Blindness The inability to see or the loss or absence of perception of visual stimuli. This condition may be the result of eye diseases; optic nerve diseases; optic chiasm diseases; or brain diseases affecting the visual pathways or occipital lobe. Retinopathy of Prematurity/visual loss | Blindness Blindness The inability to see or the loss or absence of perception of visual stimuli. This condition may be the result of eye diseases; optic nerve diseases; optic chiasm diseases; or brain diseases affecting the visual pathways or occipital lobe. Retinopathy of Prematurity in affected eye | Optic neuritis Optic neuritis Inflammation of the optic nerve. Commonly associated conditions include autoimmune disorders such as multiple sclerosis, infections, and granulomatous diseases. Clinical features include retro-orbital pain that is aggravated by eye movement, loss of color vision, and contrast sensitivity that may progress to severe visual loss, an afferent pupillary defect (Marcus-Gunn pupil), and in some instances optic disc hyperemia and swelling. Inflammation may occur in the portion of the nerve within the globe (neuropapillitis or anterior optic neuritis) or the portion behind the globe (retrobulbar neuritis or posterior optic neuritis). Cranial Nerve Palsies, retinal artery occlusion Retinal Artery Occlusion Retinal Vessel Occlusion, optic atrophy Atrophy Decrease in the size of a cell, tissue, organ, or multiple organs, associated with a variety of pathological conditions such as abnormal cellular changes, ischemia, malnutrition, or hormonal changes. Cellular Adaptation, trauma |
| OC (middle lesion) | Bitemporal heteronymous hemianopsia | Loss of temporal visual fields on both eyes Both Eyes Refractive Errors | Pituitary Pituitary A small, unpaired gland situated in the sella turcica. It is connected to the hypothalamus by a short stalk which is called the infundibulum. Hormones: Overview and Types tumor Tumor Inflammation, craniopharyngioma Craniopharyngioma Craniopharyngiomas are rare squamous epithelial tumors with a solid and/or cystic structure that arise from the remnants of Rathke’s pouch along the pituitary stalk, in the suprasellar region. Craniopharyngiomas are histologically benign but tend to invade surrounding structures; thus, they should be treated as low-grade malignancies. Craniopharyngioma, meningioma Meningioma Meningiomas are slow-growing tumors that arise from the meninges of the brain and spinal cord. The vast majority are benign. These tumors commonly occur in individuals with a history of high doses of skull radiation, head trauma, and neurofibromatosis 2. Meningioma |
| Partial optic nerve Optic nerve The 2nd cranial nerve which conveys visual information from the retina to the brain. The nerve carries the axons of the retinal ganglion cells which sort at the optic chiasm and continue via the optic tracts to the brain. The largest projection is to the lateral geniculate nuclei; other targets include the superior colliculi and the suprachiasmatic nuclei. Though known as the second cranial nerve, it is considered part of the central nervous system. The 12 Cranial Nerves: Overview and Functions lesion | Ipsilateral hemianopsia | Loss of visual field on affected eye (temporal nerve injury Nerve Injury Surgical Complications: nasal hemianopsia; nasal nerve injury Nerve Injury Surgical Complications: temporal hemianopsia) | Aneurysm Aneurysm An aneurysm is a bulging, weakened area of a blood vessel that causes an abnormal widening of its diameter > 1.5 times the size of the native vessel. Aneurysms occur more often in arteries than in veins and are at risk of dissection and rupture, which can be life-threatening. Thoracic Aortic Aneurysms, trauma |
| Optic tract | Contralateral homonymous hemianopsia | Loss of half of the visual fields opposite the lesion | Brain Brain The part of central nervous system that is contained within the skull (cranium). Arising from the neural tube, the embryonic brain is comprised of three major parts including prosencephalon (the forebrain); mesencephalon (the midbrain); and rhombencephalon (the hindbrain). The developed brain consists of cerebrum; cerebellum; and other structures in the brain stem. Nervous System: Anatomy, Structure, and Classification tumor Tumor Inflammation, abscess Abscess Accumulation of purulent material in tissues, organs, or circumscribed spaces, usually associated with signs of infection. Chronic Granulomatous Disease ( temporal lobe Temporal lobe Lower lateral part of the cerebral hemisphere responsible for auditory, olfactory, and semantic processing. It is located inferior to the lateral fissure and anterior to the occipital lobe. Cerebral Cortex: Anatomy), middle cerebral artery Middle cerebral artery The largest of the cerebral arteries. It trifurcates into temporal, frontal, and parietal branches supplying blood to most of the parenchyma of these lobes in the cerebral cortex. These are the areas involved in motor, sensory, and speech activities. Cerebrovascular System: Anatomy infarction |
| Optic radiation Radiation Emission or propagation of acoustic waves (sound), electromagnetic energy waves (such as light; radio waves; gamma rays; or x-rays), or a stream of subatomic particles (such as electrons; neutrons; protons; or alpha particles). Osteosarcoma (Meyer’s loop or inferior radiation Radiation Emission or propagation of acoustic waves (sound), electromagnetic energy waves (such as light; radio waves; gamma rays; or x-rays), or a stream of subatomic particles (such as electrons; neutrons; protons; or alpha particles). Osteosarcoma/lateral bundle) | Contralateral homonymous superior quadrantanopsia | Loss of superior quadrant visual fields opposite the lesion (“pie in the sky”) | Tumor Tumor Inflammation ( occipital Occipital Part of the back and base of the cranium that encloses the foramen magnum. Skull: Anatomy, temporal lobe Temporal lobe Lower lateral part of the cerebral hemisphere responsible for auditory, olfactory, and semantic processing. It is located inferior to the lateral fissure and anterior to the occipital lobe. Cerebral Cortex: Anatomy) |
| Optic radiation Radiation Emission or propagation of acoustic waves (sound), electromagnetic energy waves (such as light; radio waves; gamma rays; or x-rays), or a stream of subatomic particles (such as electrons; neutrons; protons; or alpha particles). Osteosarcoma (superior radiation Radiation Emission or propagation of acoustic waves (sound), electromagnetic energy waves (such as light; radio waves; gamma rays; or x-rays), or a stream of subatomic particles (such as electrons; neutrons; protons; or alpha particles). Osteosarcoma/medial bundles) | Contralateral homonymous inferior quadrantanopsia | Loss of inferior quadrant visual fields opposite the lesion (“pie on the floor”) | Tumor Tumor Inflammation ( occipital Occipital Part of the back and base of the cranium that encloses the foramen magnum. Skull: Anatomy, parietal lobe Parietal lobe Upper central part of the cerebral hemisphere. It is located posterior to central sulcus, anterior to the occipital lobe, and superior to the temporal lobes. Cerebral Cortex: Anatomy) |
| Optic radiation Radiation Emission or propagation of acoustic waves (sound), electromagnetic energy waves (such as light; radio waves; gamma rays; or x-rays), or a stream of subatomic particles (such as electrons; neutrons; protons; or alpha particles). Osteosarcoma | Contralateral homonymous hemianopsia | Loss of half of the visual fields opposite the lesion | Tumor Tumor Inflammation, middle cerebral artery Middle cerebral artery The largest of the cerebral arteries. It trifurcates into temporal, frontal, and parietal branches supplying blood to most of the parenchyma of these lobes in the cerebral cortex. These are the areas involved in motor, sensory, and speech activities. Cerebrovascular System: Anatomy infarction |
| Primary visual cortex Visual cortex Area of the occipital lobe concerned with the processing of visual information relayed via visual pathways. Cerebral Cortex: Anatomy | Contralateral homonymous hemianopsia with macular sparing | Loss of half of the visual fields opposite the lesion, but central vision Vision Ophthalmic Exam is maintained due to collateral circulation Circulation The movement of the blood as it is pumped through the cardiovascular system. ABCDE Assessment ( macula Macula An oval area in the retina, 3 to 5 mm in diameter, usually located temporal to the posterior pole of the eye and slightly below the level of the optic disk. It is characterized by the presence of a yellow pigment diffusely permeating the inner layers, contains the fovea centralis in its center, and provides the best phototropic visual acuity. It is devoid of retinal blood vessels, except in its periphery, and receives nourishment from the choriocapillaris of the choroid. Eye: Anatomy receives blood supply from the middle cerebral artery Middle cerebral artery The largest of the cerebral arteries. It trifurcates into temporal, frontal, and parietal branches supplying blood to most of the parenchyma of these lobes in the cerebral cortex. These are the areas involved in motor, sensory, and speech activities. Cerebrovascular System: Anatomy) | Posterior cerebral artery Posterior cerebral artery Artery formed by the bifurcation of the basilar artery. Branches of the posterior cerebral artery supply portions of the occipital lobe; parietal lobe; inferior temporal gyrus, brainstem, and choroid plexus. Cerebrovascular System: Anatomy thrombosis Thrombosis Formation and development of a thrombus or blood clot in the blood vessel. Epidemic Typhus, trauma, tumor Tumor Inflammation |

Visual field defects:
1. injury to the right (R) macula: right (R) central scotoma
2. injury to the R optic nerve: R visual loss
3. injury to the OC: bitemporal hemianopsia
4. injury to the R temporal optic nerve: R nasal hemianopsia
5. injury to the R optic tract: left (L) homonymous hemianopsia
6. injury to the R Meyer’s loop: L homonymous superior quadrantanopsia
7. injury to the R superior optic radiation: L homonymous inferior quadrantanopsia
8. injury to the R optic radiation: L homonymous hemianopsia
9. injury to the R primary visual cortex: L homonymous hemianopsia with macular sparing due to collateral blood supply