LosLOSNeisseria receptores son proteínas ubicadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la superficie o dentro de una célula que pueden unirse a moléculas de señalización conocidas como ligandos (e.g., hormonas) y provocar algún tipo de respuesta dentro de la célula. LosLOSNeisseria receptores de superficie están unidos a la membrana celular, reciben señales de su entorno y las transmiten a la célula, a menudo a través de la generación de 2dos mensajeros (como el adenosín monofosfato cíclico) o mediante cascadas de fosforilación. Hay múltiples subclases diferentes de receptores de superficie, y 3 de las clases más importantes incluyen receptores de canales iónicos activados por ligando, receptores ligados a enzimas (losLOSNeisseria más comunes son losLOSNeisseria receptores de tirosina quinasas) y receptores acoplados a proteína G. LosLOSNeisseria receptores intracelulares, por otro lado, están ubicados dentro del citoplasma y, a menudo, actúan como factores de transcripción, interactuando directamente con el ácido desoxirribonucleico (ADN) y afectando la expresión génica.
LosLOSNeisseria receptores son proteínas ubicadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la superficie o dentro de una célula que pueden unirse a moléculas de señalización conocidas como ligandos (e.g., hormonas) y provocar algún tipo de respuesta dentro de la célula.
Fisiología general
El ligando se une alALAmyloidosisreceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors → induce un cambio conformacional enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la proteína receptora
El ligando puede provenir de:
Contacto directo, a través de uniones estrechas
Señalización endocrina y paracrina (a través de hormonas)
Señalización sináptica (a través de neurotransmisores)
La vía de transducción de señales dentro de la célula suele ser un proceso complejo de varios pasos.
LosLOSNeisseria resultados finales pueden incluir:
Alteración de la expresión génica (↑ o ↓ producción de proteínas específicas)
Apertura y/o cierre de canales de membrana
Liberación de productos celulares alALAmyloidosis líquido extracelular o alALAmyloidosis torrente sanguíneo
Clasificación: Receptores de Superficie Celular versus Intracelulares
LosLOSNeisseria receptores se pueden dividir enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum 2 categorías principales: receptores de superficie celular (transmembrana) y receptoresintracelulares.
Receptores de superficie celular (receptores transmembrana)
Situados dentro de la membrana plasmática
Constan de 3 dominios:
Dominio de unión a ligando extracelular
Dominio hidrofóbico dentro de la membrana plasmática
Dominio intracelular
LosLOSNeisseria ligandos que utilizan receptores transmembrana normalmente no pueden atravesar la propia membrana porque son hidrófilos y/o grandes.
El dominio intracelular comunica la señal dentro de la célula a través de:
Modificación covalente de otras moléculas:
Generalmente, a través de la fosforilación, lo que desencadena una cascada de fosforilación
Cascada de fosforilación: una secuencia de varios pasos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la que un grupo fosfato pasa de 1 molécula a la siguiente mediante una serie de enzimas conocidas como quinasas
Generación de 2dos mensajeros, siendo losLOSNeisseria más comunes:
Adenosín monofosfato cíclico
Guanosín monofosfato cíclico
Trifosfato de inositol
Diacilglicerol
Iones de calcio (CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+)
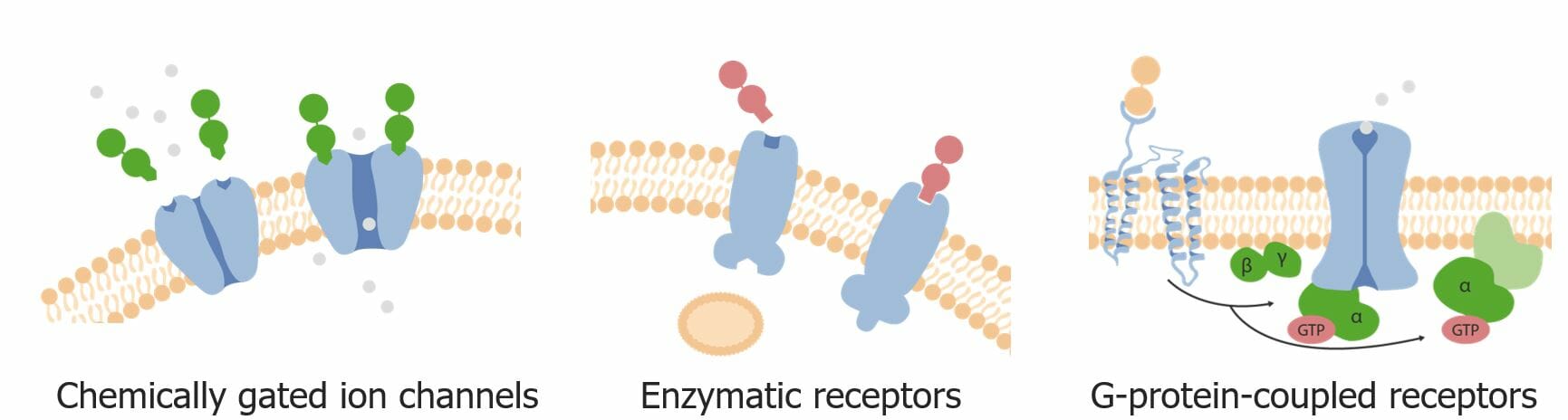
Subtipos de receptores de superficie celular:
Receptores de canales iónicos activados por ligandos (activados químicamente)
Receptores acoplados a proteína G
Receptores ligados a enzimas (tipo más común: receptores tirosina quinasas)
Tres tipos principales de receptores de superficie celular (i.e., transmembrana): canales iónicos activados químicamente, receptores enzimáticos y receptores acoplados a proteína G
Imagen por Lecturio.
Receptores intracelulares
Localizados dentro de la célula; pueden estar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum:
Dominio que interactúa con otros factores de transcripción (e.g., coactivadores, inhibidores)
LosLOSNeisseria ligandos suelen ser pequeñas moléculas hidrofóbicas que pueden atravesar la membrana celular.
Ejemplos de receptores intracelulares:
Receptores nucleares de hormonas esteroideas
ReceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors de vitamina D
Guanilato ciclasa (el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors de óxido nítrico que funciona como una enzima, generando guanosín monofosfato cíclico)
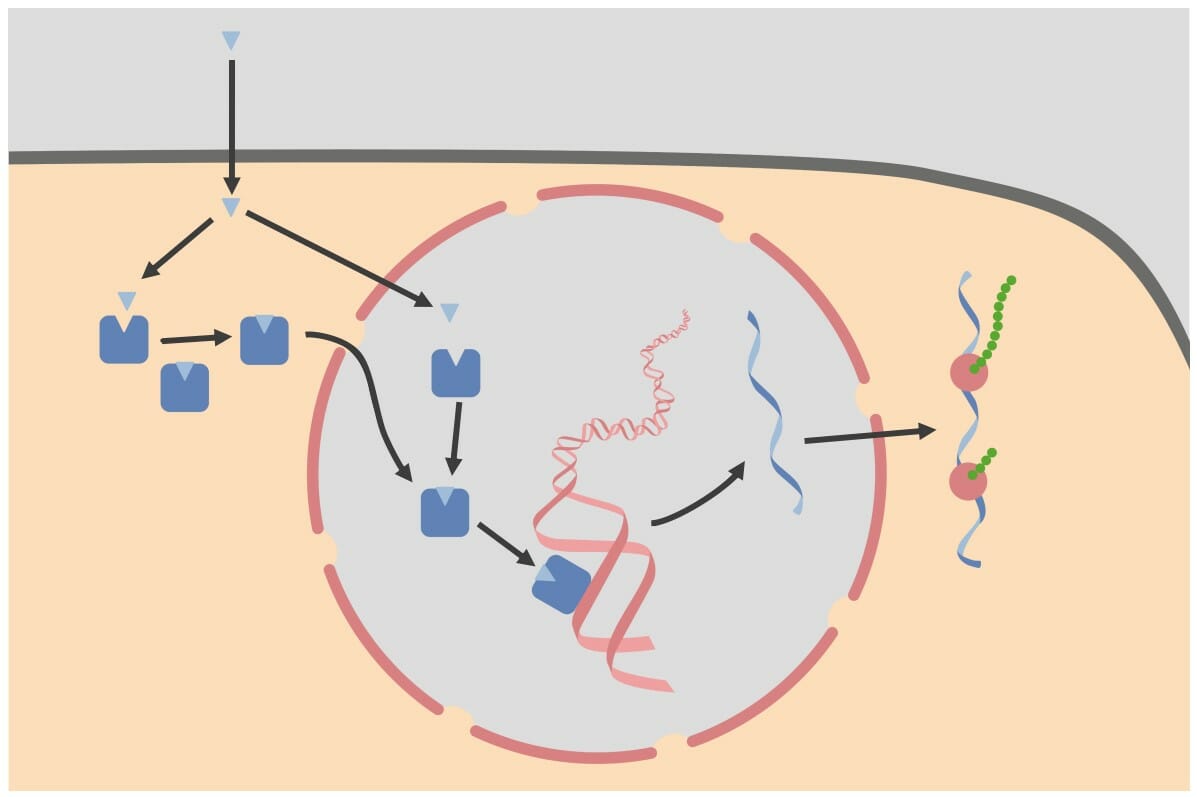
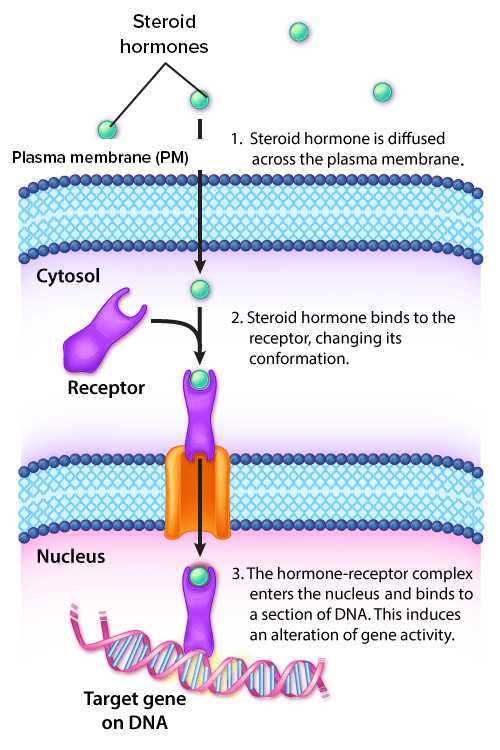
Receptores intracelulares: Los receptores intracelulares pueden estar ubicados dentro del citoplasma o del núcleo. Los receptores ubicados dentro del citoplasma se mueven hacia el núcleo una vez que se unen con su ligando (i.e., hormona). Dentro del núcleo, el complejo hormona-receptor se une a los elementos de respuesta hormonal, que son secuencias específicas de ADN. El complejo luego se une a otros factores de transcripción para alterar la expresión génica.
Imagen por Lecturio.
Tabla: Tipos de receptores y ligandos
Tipos de receptores
Ejemplo de ligandos
Receptores intracelulares
Receptores nucleares
Hormonas esteroideas (e.g., glucocorticoides)
Receptores de superficie celular
Receptores acoplados a proteína G
Catecolaminas
ReceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasas
Receptores de Canales Iónicos Activados por Ligandos
LosLOSNeisseria receptores de canales iónicos activados por ligandos (activados químicamente) son un subtipo de receptores de superficie celular.
Estructura
Constan de múltiples subunidades transmembrana alrededor de un canal central conductor de iones
Tienen un sitio de unión alALAmyloidosis ligando extracelular (o intracelular)
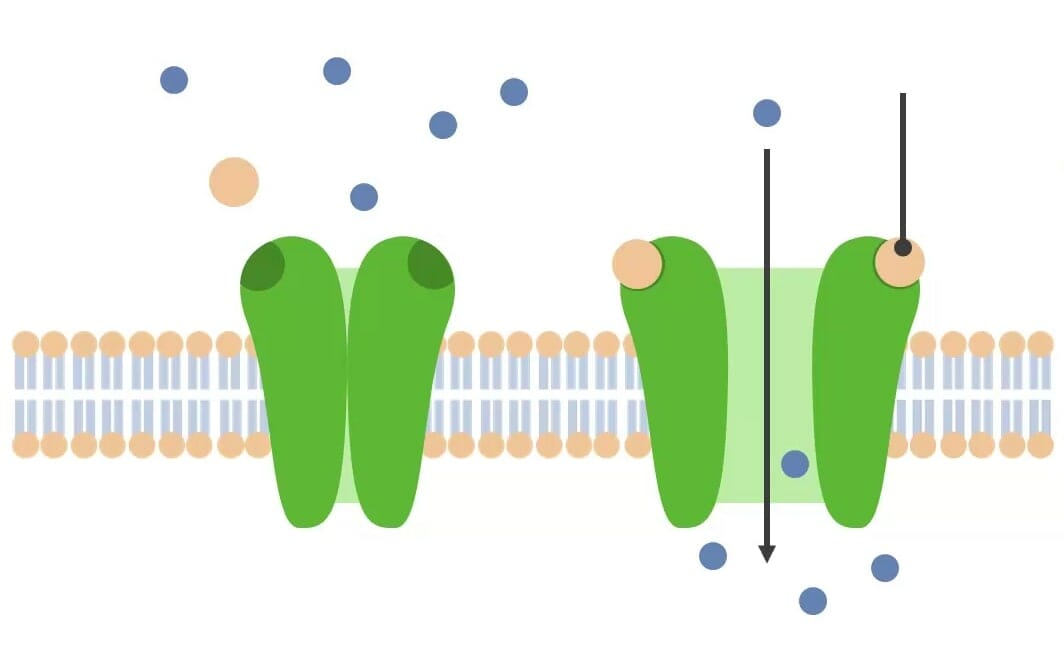
Canales iónicos activados por ligandos (activados químicamente): Esta imagen muestra la unión de un ligando (bola marrón) con un receptor de canal iónico activado por ligando (estructura verde), lo que provoca un cambio de conformación en el canal, lo que permite el paso de iones.
Imagen por Lecturio.
Fisiología
El ligando (e.g., acetilcolina) se une alALAmyloidosisreceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors del canal iónico.
Induce un cambio conformacional → se abre el canal
LosLOSNeisseria iones fluyen siguiendo su gradiente electroquímico a través de losLOSNeisseria canales (e.g., Na+, K+, CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ y/o Cl−).
Esto puede resultar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum:
Transmisión de señales nerviosas
Contracción muscular
Liberación de hormonas
Activación de células T
Cuando el ligando se disocia → el canal se cierra
Ejemplo de relevancia clínica
Miastenia gravis: un trastorno neuromuscular autoinmune caracterizado por debilidad y fatigabilidad de losLOSNeisseria músculos esqueléticos causados por la disfunción/destrucción de losLOSNeisseria receptores de acetilcolina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la unión neuromuscular (un tipo de receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors de canal iónico activado por ligando). Cuando la acetilcolina se une, losLOSNeisseria canales se abren, lo que permite la entrada de Na+enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la célula, lo que da como resultado una despolarización que finalmente conduce a la contracción muscular. Sin losLOSNeisseria receptores de acetilcolina normales, la contracción muscular es anormal, por lo que la miastenia gravis se presenta con fatiga, ptosisPtosisCranial Nerve Palsies, disfagia, dificultades respiratorias y debilidad progresiva enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las extremidades, lo que provoca dificultad enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el movimiento.
Receptores Ligados a Enzimas
Descripción general
Receptores que tienen algún tipo de actividad enzimática cuando son “activados” por su ligando
Frecuentemente quinasas: enzimas que catalizan la transferencia de un grupo fosfato de 1 molécula a otra
Tirosina quinasas:
Tipo más común
Se han identificado más de 90 genesGenesA category of nucleic acid sequences that function as units of heredity and which code for the basic instructions for the development, reproduction, and maintenance of organisms.DNA Types and Structure que codifican para receptores tirosina quinasas
Serina-treonina quinasas
Receptores asociados a quinasas: losLOSNeisseria receptores no poseen actividad quinasa intrínseca por sí mismos, pero se unen a quinasas cuando se activan.
LosLOSNeisseria receptores tirosina quinasas están involucrados enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum una serie de procesos frecuentemente asociados con la función celular general:
Ciclo celular
Crecimiento y proliferación celular
Migración celular
Metabolismo celular
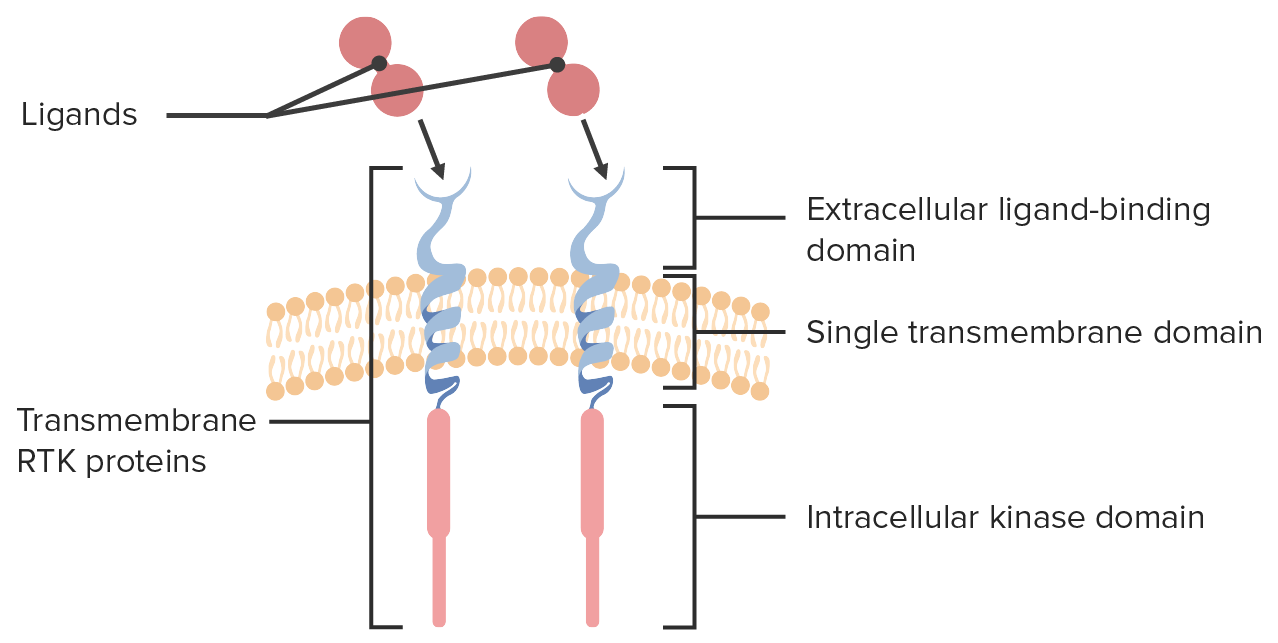
Estructura del receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasa
Dominio extracelular que contiene el sitio de unión alALAmyloidosis ligando
Dominio de quinasa intracelular
Hélice transmembrana única que une componentes extracelulares e intracelulares
La estructura de 3 partes del receptor tirosina quinasa se muestra junto con los ligandos que se unen a sus sitios de unión extracelulares.
Imagen por Lecturio.
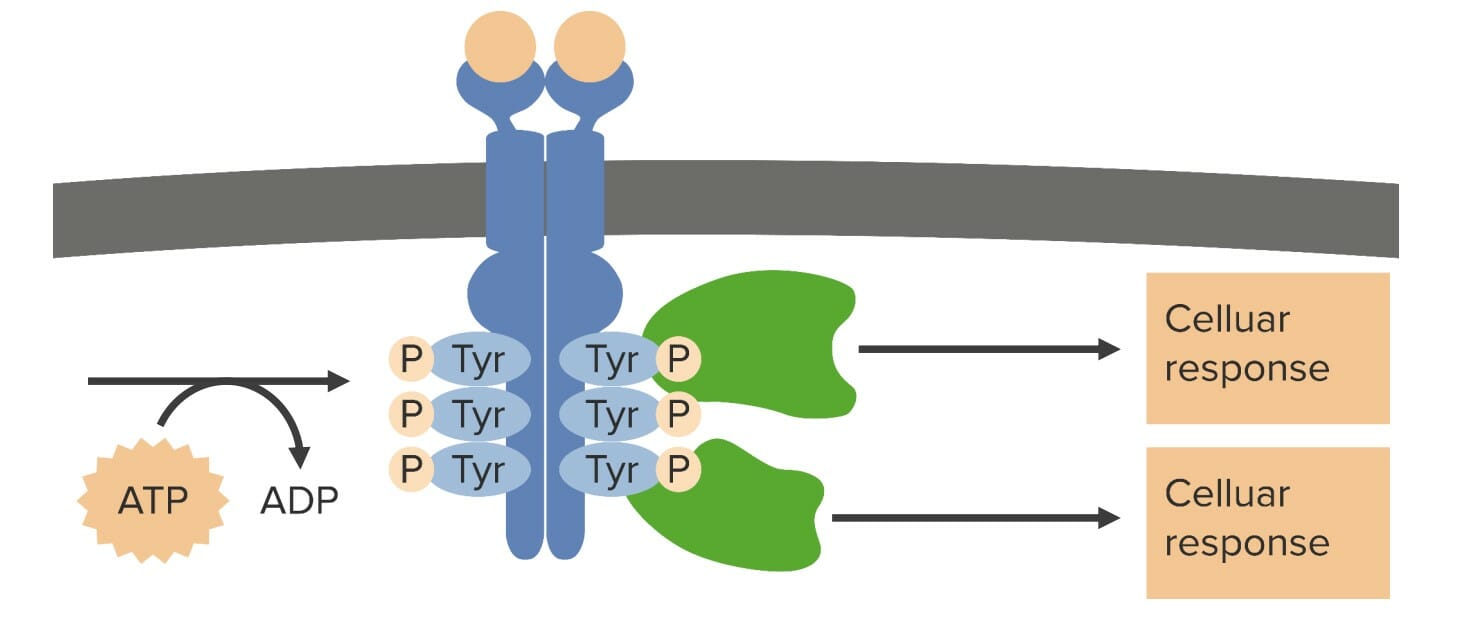
Vía de señalización del receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasa
El ligando se une a losLOSNeisseria receptores tirosina quinasa individuales, lo que conduce a:
Dimerización: 2 receptores tirosina quinasa vecinos se unen.
1 monómero de receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasa transfiere un grupo fosfato del adenosín trifosfato alALAmyloidosis otro monómero de receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasa (ahora conocido como fosfotirosina).
Las fosfotirosinas actúan como sitios de acoplamiento para otras proteínas involucradas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la cascada de transducción de señales.
La respuesta celular de la activación del receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors tirosina quinasa depende de las proteínas subsecuentes enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el proceso que están presentes.
La ventaja de una cascada de quinasas es que una pequeña señal externa puede amplificarse muchas veces dentro de la célula.
La disfunción de esta cascada puede provocar cáncer debido a la falta de control de la función celular general.
Función del receptor tirosina quinasa: Cuando un ligando se une al dominio extracelular del receptor tirosina quinasa, 2 receptores tirosina quinasa se unen en un proceso conocido como dimerización. Una vez dimerizado, cada uno de los monómeros transfiere un grupo fosfato del adenosín trifosfato (ATP) a su compañero en un proceso conocido como autofosforilación. Después de la autofosforilación, las fosfotirosinas actúan como sitios de acoplamiento y activación para otras proteínas, que suelen ser otras enzimas. El tipo de respuesta celular depende de qué proteínas adicionales estén presentes. Tyr: tirosina P: fosfato
Imagen por Lecturio.
Ejemplos de receptor tirosina quinasa
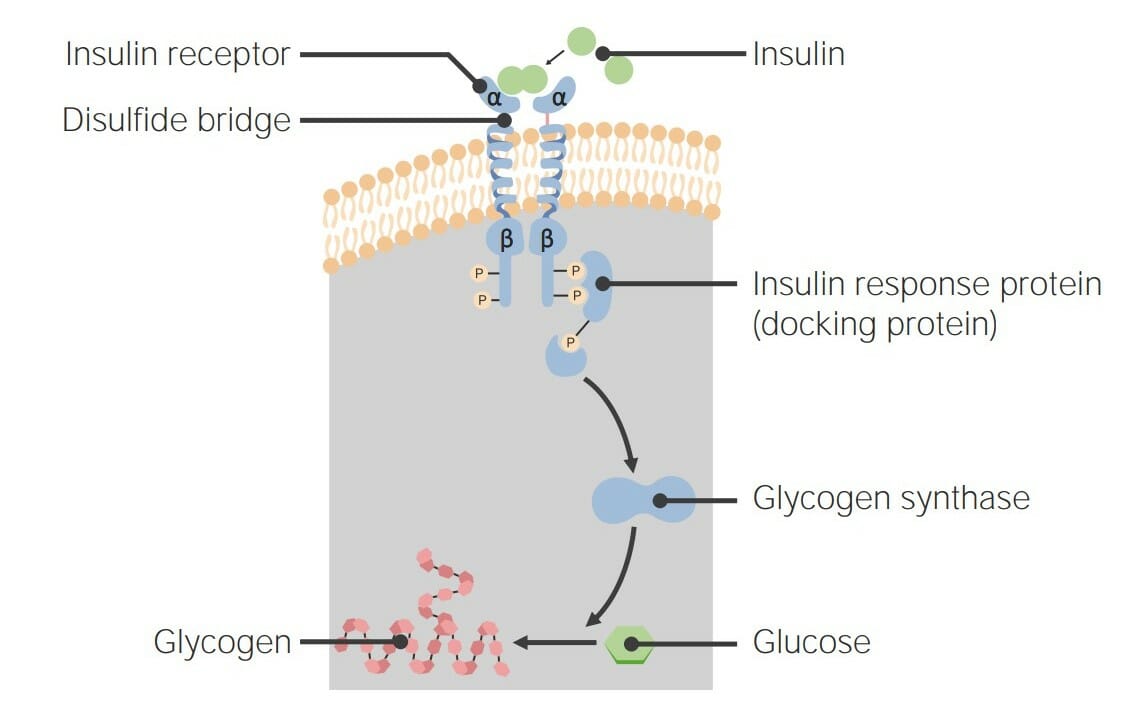
Receptores de insulina:
LosLOSNeisseria receptores de insulina maduros comienzan como un dímero inactivo, que contiene 2 subunidades alfa y 2 beta que están unidas entre sí por puentes disulfuro:
Subunidades alfa: dominios extracelulares que contienen el sitio de unión alALAmyloidosis ligando
Subunidades beta: dominios de tirosina quinasa transmembrana e intracelular
La insulina se une a losLOSNeisseria dominios alfa → desencadena una autofosforilación rápida
Vía de señalización:
El sustrato de respuesta a la insulina 1 se une a las fosfotirosinas →
Activa la fosfatidilinositol quinasa →
Resultando enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la formación de fosfatidilinositol (3,4,5)-trifosfato →
El fosfatidilinositol (3,4,5)-trifosfato activa la proteína quinasa 1 dependiente de 3-fosfoinosítidos, que activa la quinasa Akt →
Causa la translocación del transportador de glucosa 4 (un canal de proteína que permite que la glucosa ingrese a la célula) a la membrana plasmática →
↑ La glucosa que puede entrar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la célula
Otra vía de señalización de la insulina da como resultado una mayor transcripción y traducción de la glucógeno-sintasa → ↑ conversión de glucosa enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum glucógeno dentro de la célula
El receptor de insulina, un receptor de tirosina quinasa: Cuando la insulina se une, la proteína de respuesta a la insulina desencadena una cascada de fosforilación, lo que finalmente resulta en la activación de la glucógeno-sintasa, que convierte la glucosa extra en glucógeno.
Imagen por Lecturio.
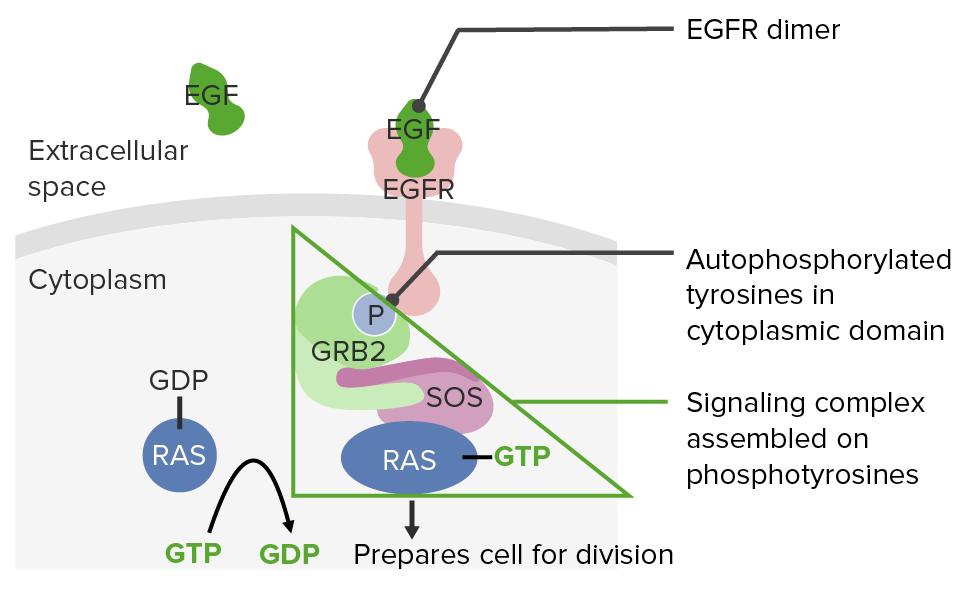
Receptores del factor de crecimiento epidérmico y la vía de señalización RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis:
El factor de crecimiento epidérmico se une a sus receptores → provocando dimerización y autofosforilación
Un complejo de señalización de proteínas se ensambla enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las fosfotirosinas.
Este complejo de señalización transfiere un grupo fosfato del guanosín trifosfato alALAmyloidosis difosfato de guanosina RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis (RAS-GDP), lo que da como resultado RAS-GTP, que es la forma activa de RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis.
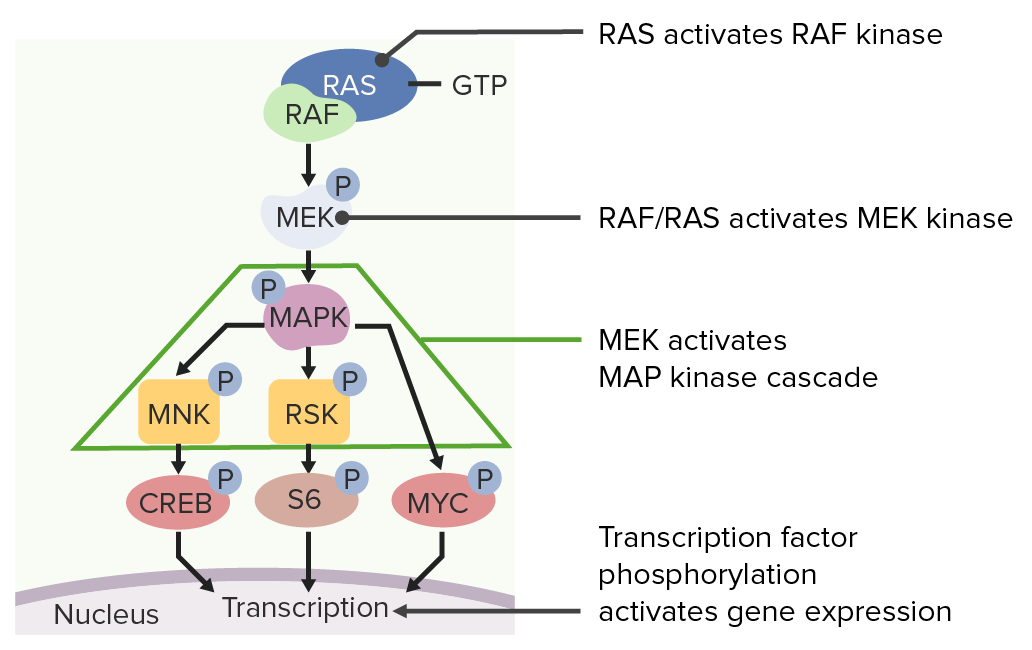
Vía de señalización RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis:
MAPK activa MNK y RSK, que fosforilan factores de transcripción específicos necesarios para la división celular.
RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery StenosisenENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum sí mismo es una GTPasa lenta:
RAS-GTP escindirá lentamente un grupo fosfato, inactivándose de nuevo a RAS-GDP.
Mutaciones puntuales pueden remover la actividad GTPasa, resultado enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum un RASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis activado permanentemente → promueve la división celular continua
Las mutaciones enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumRASRASRenal artery stenosis (RAS) is the narrowing of one or both renal arteries, usually caused by atherosclerotic disease or by fibromuscular dysplasia. If the stenosis is severe enough, the stenosis causes decreased renal blood flow, which activates the renin-angiotensin-aldosterone system (RAAS) and leads to renovascular hypertension (RVH).Renal Artery Stenosis están involucradas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum varios tipos de cánceres enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum humanos.
El factor de crecimiento epidérmico (EGF) se une al receptor de EGF (EGFR), lo que resulta en la fosforilación de RAS.
Imagen por Lecturio.
El RAS fosforilado luego activa una cascada de señalización que da como resultado la división celular.
Imagen por Lecturio.
Ejemplos de relevancia clínica
Se sabe que las anomalías en los receptores tirosina quinasa causan varios síndromes de malformaciones congénitas y cánceres diferentes, especialmente con mutaciones de ganancia de función que causan una división celular excesiva. Algunos ejemplos son:
Leucemia mieloide crónica: una proliferación maligna de la línea celular granulocítica debido a una translocación recíproca (9;22)(q34;q11). El cromosoma contiene un gen de fusión BCR-ABL1 (de ABL1enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el cromosoma 9 y BCRBCRLymphocytes: HistologyenENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el cromosoma 22), que produce tirosina cinasas constitutivamente activas y, enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum consecuencia, una producción granulocítica descontrolada. LosLOSNeisseria pacientes pueden estar asintomáticos o tener síntomas constitucionales, dolorDolorInflammation esternal y esplenomegalia.
Acondroplasia: una displasia ósea autosómica dominante debida a mutaciones de ganancia de función en el gen del receptor 3 del factor de crecimiento de fibroblastos, que codifica un receptor tirosina quinasa. Como resultado, el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors 3 del factor de crecimiento de fibroblastos se activa permanentemente, lo que inhibe la proliferación de condrocitos, lo que da como resultado una formación ósea deficiente y anomalías esqueléticas.
LosLOSNeisseria receptores acoplados a proteína G son proteínas transmembrana que se unen a ligandos hormonales enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum su lado extracelular, lo que induce un cambio conformacional dentro de la célula, activando una proteína G asociada, que luego desencadena una cascada de señalización a través de 2dos mensajeros.
La familia más grande de receptores, con más de 800 genesGenesA category of nucleic acid sequences that function as units of heredity and which code for the basic instructions for the development, reproduction, and maintenance of organisms.DNA Types and Structure de receptores acoplados a proteína G identificados
A menudo involucrados enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum cambios metabólicos y estructurales dentro de una célula
Ancla el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.ReceptorsenENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana plasmática
A menudo compuesto por 7 hélices alfa (conocidas como receptores de siete transmembranas)
Dominio intracelular: unido a una proteína G
Proteínas G:
Proteínas que se unen a losLOSNeisseria nucleótidos de guanina (guanosín trifosfato y guanosín difosfato)
Consta de 3 subunidades: alfa, beta y gamma
Subunidad alfa:
Se une a losLOSNeisseria nucleótidos de guanina: guanosín trifosfato (estado activo) y guanosín difosfato (estado inactivo)
Lentamente, hidroliza guanosín trifosfato a guanosín difosfato → una vez activada por guanosín trifosfato, la subunidad alfa eventualmente se inactivará a través de la conversión hidrolítica a guanosín difosfato
Beta y gamma: ayudan alALAmyloidosis complejo alfa‒guanosín difosfato a asociarse con el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G
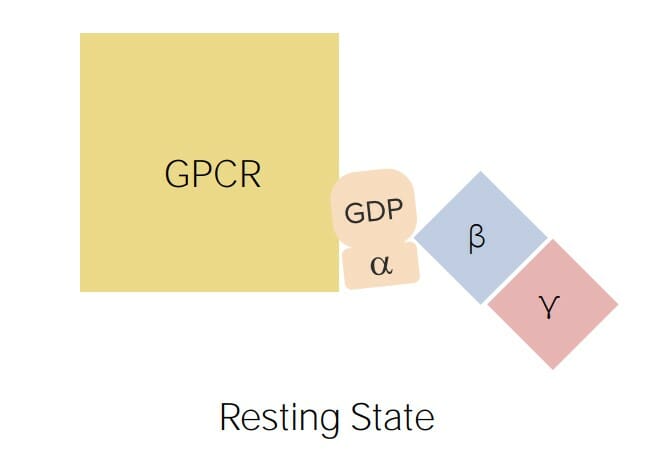
Diagrama que representa un receptor acoplado a proteína G (GPCR) unido a una proteína G: Las proteínas G constan de 3 subunidades: alfa (que se une al guanosín difosfato (GDP) en su forma inactiva y al guanosín trifosfato (GTP) en su forma activa), beta y gamma (que ayuda a la subunidad alfa a asociarse con el GPCR).
Imagen por Lecturio.
Vía de activación
El ligando se une a su sitio de unión enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la porción externa del receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors → induce un cambio conformacional enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G
Esto provoca la fosforilación del guanosín difosfato unido a la subunidad alfa → se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum alfa‒guanosín trifosfato (proteína G activada)
Alfa‒guanosín trifosfato se separa de la subunidad beta y gamma → fosforila la siguiente proteína enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la cascada de señalización conocida como proteína efectora
Proteínas efectoras:
Enzimas, a menudo unidas a la membrana
La fosforilación puede ser activadora o inhibidora
Genera con frecuencia 2dos mensajeros
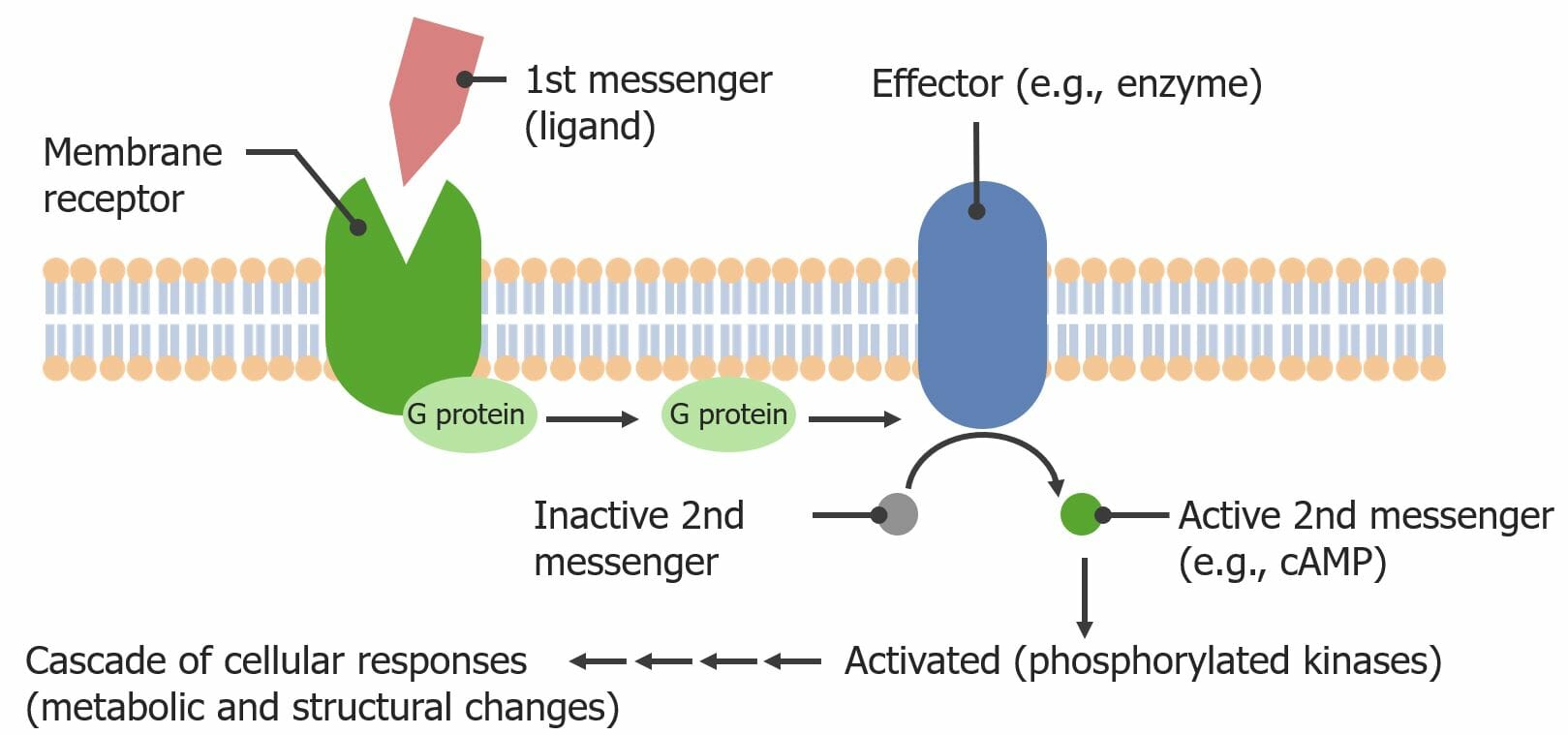
Vía de activación de los receptores acoplados a proteína G: Un ligando se une al receptor acoplado a proteína G, induciendo un cambio conformacional interno. Este cambio conformacional hace que la subunidad alfa de la proteína G intercambie un guanosín difosfato (GDP) por un guanosín trifosfato (GTP), que activa la proteína G. La subunidad alfa unida a GTP se separa de las subunidades beta y gamma y activa una enzima efectora (a través de la fosforilación del GTP). La enzima efectora luego activa un 2do mensajero (aquí, el adenosín monofosfato cíclico (cAMP)), que transmite la señal dentro de la célula.
Imagen por Lecturio.
Adenilato ciclasa y el sistema de 2do mensajero adenosín monofosfato cíclico
La adenilato ciclasa (también llamada adenilil ciclasa) es una proteína efectora común de losLOSNeisseria receptores acoplados a proteína G. Una proteína G activada fosforila la adenilato ciclasa, activándola para convertir adenosín trifosfato enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum adenosín monofosfato cíclico, un 2do mensajero común.
Ejemplo 1: vía del glucagón
El glucagón se une a su receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors, que es un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G → cambio conformacional
La subunidad alfa de la proteína G unida a la superficie interna del receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G intercambia guanosín difosfato por guanosín trifosfato y se separa de las subunidades beta y gamma.
Alfa‒guanosín trifosfato (forma activa) fosforila a la adenilato ciclasa, activándola.
La adenilato ciclasa genera adenosín monofosfato cíclico a partir de adenosín trifosfato.
El adenosín monofosfato cíclico fosforila las subunidades reguladoras de la proteína quinasa A → esto haceHACEAltitude Sickness que las subunidades reguladoras se disocien de las subunidades catalíticas, “activando” así la proteína quinasa A
La proteína quinasa A fosforila:
Glucógeno-sintasa, inactivándola → se detiene la síntesis de glucógeno
Fosforilasa quinasa, activándola → fosforilasa quinasa activa la glucógeno-fosforilasa → estimula la descomposición del glucógeno
Resumen: el glucagón estimula la descomposición del glucógeno enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum glucosa y simultáneamente inhibe la síntesis de glucógeno.
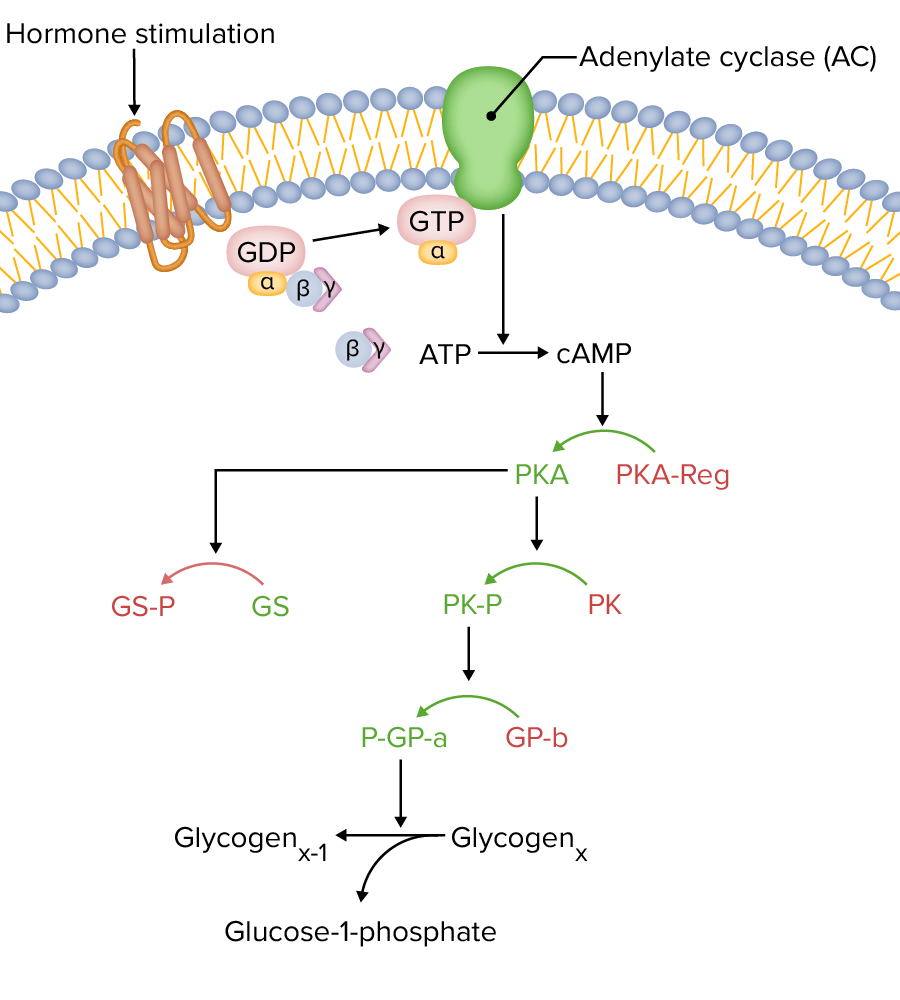
Receptor acoplado a proteína G acoplado a adenilato ciclasa (AC):
Nota: las enzimas activas se muestran en verde, mientras que las inactivas se muestran en rojo. La AC convierte el adenosín trifosfato (ATP) en adenosín monofosfato cíclico (cAMP), que luego activa la proteína quinasa A (PKA). Luego, la PKA fosforila tanto la glucógeno-sintasa (GS), inactivándola, como la fosforilasa quinasa (PK), activándola. La PK activada luego activa la glucógeno-fosforilasa, que estimula la descomposición del glucógeno en glucosa.
GTP: guanosín-5′-trifosfato
GDP: guanosín difosfato
ATP: adenosín trifosfato
cAMP: adenosín monofosfato cíclico
Imagen por Lecturio.
Ejemplo 2: Vías de epinefrina (1 molécula que tiene diferentes efectos dependiendo de las proteínas presentes enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la célula diana)
EnENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el músculo esquelético: la epinefrina se une a un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G, que activa la adenilato ciclasa de forma idéntica alALAmyloidosis glucagón → ↑ descomposición del glucógeno enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum glucosa
EnENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el músculo cardíaco: el adenosín monofosfato cíclico producido por la adenilato ciclasa resulta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum ↑ fuerza de contracción
EnENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria intestinos: la epinefrina se une a un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G que inhibe la adenilato ciclasa, lo que provoca la relajación de losLOSNeisseria músculos intestinales y la ralentización de la digestión
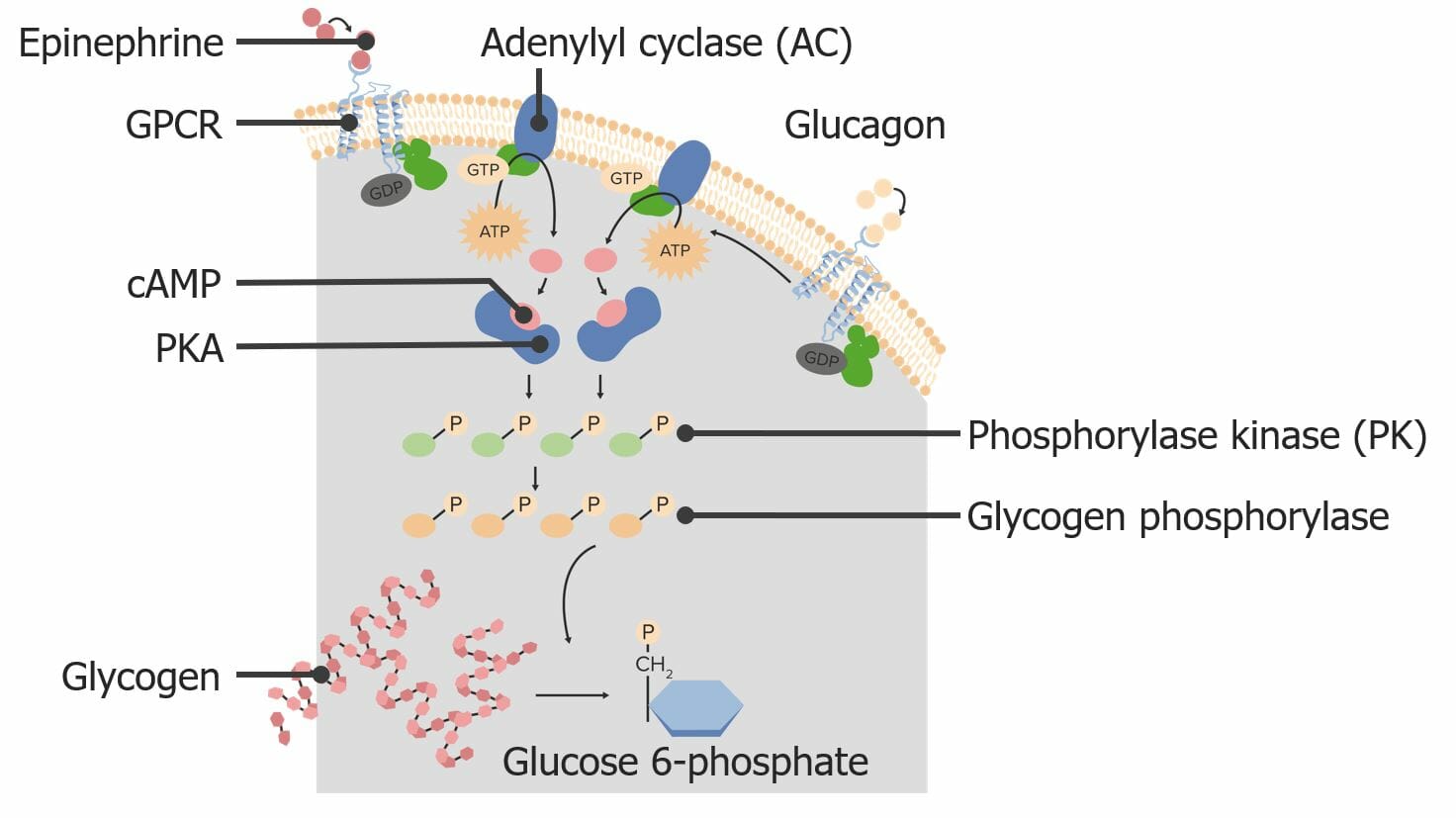
Dos proteínas G diferentes pueden activar la misma vía de transducción de señales internas: Aquí, la epinefrina y el glucagón pueden activar la descomposición del glucógeno en glucosa. GPCR: receptores acoplados a proteína G cAMP: adenosín monofosfato cíclico PKA: proteína quinasa A
Imagen por Lecturio.
Fosfolipasa C (proteína efectora) y 2dos mensajeros trifosfato de inositol y diacilglicerol
El ligando se une alALAmyloidosisreceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G → cambio conformacional
La subunidad alfa de la proteína G intercambia guanosín difosfato por guanosín trifosfato y se separa de las subunidades beta y gamma.
Alfa‒guanosín trifosfato (activo) fosforila la fosfolipasa C, activándola.
La fosfolipasa C divide el fosfatidilinositol 4,5-bisfosfato enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum trifosfato de inositol y diacilglicerol.
El trifosfato de inositol viaja alALAmyloidosis retículo endoplasmático y se une a un canal iónico controlado por ligando, lo que haceHACEAltitude Sickness que ese canal se abra y libere CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ hacia el citoplasma.
El CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ entonces:
Ejerce una respuesta celular
HaceHACEAltitude Sickness que el diacilglicerol active la proteína quinasa C, que luego fosforila proteínas adicionales
E.g., enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células parietales del estómago, la acetilcolina se une a un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G muscarínico → activa fosfolipasa C → genera trifosfato de inositol y diacilglicerol → CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ desde el retículo endoplasmático y diacilglicerol ayudan a activar la H+/K+ ATPasa, que secreta iones H+enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la luz estomacal
Un receptor acoplado a proteína G activa la fosfolipasa C (PLC), que convierte el fosfatidilinositol 4,5-bisfosfato (PIP2) en trifosfato de inositol (IP3) y diacilglicerol (DAG). Luego, el IP3 se une a un canal iónico activado por ligando en el retículo endoplasmático (ER), lo que hace que el canal se abra y el Ca2+ fluya hacia el citoplasma, lo que provoca una respuesta celular (e.g., desencadena la liberación de hormonas de las células endocrinas). GTP: guanosín-5′-trifosfato GDP: guanosín difosfato
Imagen por Lecturio.
Ejemplo de relevancia clínica
Más de 30 enfermedades humanas diferentes pueden vincularse a mutaciones enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria receptores acoplados a proteína G. Estas mutaciones pueden ser activantes o inhibidoras. Un ejemplo es la diabetesDiabetesDiabetes mellitus (DM) is a metabolic disease characterized by hyperglycemia and dysfunction of the regulation of glucose metabolism by insulin. Type 1 DM is diagnosed mostly in children and young adults as the result of autoimmune destruction of β cells in the pancreas and the resulting lack of insulin. Type 2 DM has a significant association with obesity and is characterized by insulin resistance.Diabetes Mellitus insípida nefrogénica.
DiabetesDiabetesDiabetes mellitus (DM) is a metabolic disease characterized by hyperglycemia and dysfunction of the regulation of glucose metabolism by insulin. Type 1 DM is diagnosed mostly in children and young adults as the result of autoimmune destruction of β cells in the pancreas and the resulting lack of insulin. Type 2 DM has a significant association with obesity and is characterized by insulin resistance.Diabetes Mellitus insípida nefrogénica: un trastorno causado por anomalías enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors de la hormona antidiurética (ADH, por sus siglas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum inglés), que conduce a la resistencia a la ADH. El receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors ADH es un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G que normalmente desencadena la inserción de canales de acuaporina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las membranas de las células del conducto colector renal, lo que permite la reabsorción de agua. Con un receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors anormal, el receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors acoplado a proteína G transmite de manera ineficaz la señal de ADH dentro de la célula, lo que da como resultado una resistencia a la ADH y una disminución de la capacidad de las personas para concentrar la orina.
Un subtipo de receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors intracelular
Actúan como factores de transcripción activados por ligandos, que finalmente afectan la expresión génica
Aunque se conocen como receptores nucleares, a menudo se encuentran enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el citoplasma, y se mueven hacia el núcleo.
Dominio de unión alALAmyloidosis ADN central, que se une a losLOSNeisseria elementos de respuesta hormonal enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ADN
Regiones que interactúan con otros factores de transcripción
Vía de activación
El ligando se une alALAmyloidosisreceptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors intracelular (generalmente enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el citoplasma).
El complejo ligando–receptorReceptorReceptors are proteins located either on the surface of or within a cell that can bind to signaling molecules known as ligands (e.g., hormones) and cause some type of response within the cell.Receptors viaja alALAmyloidosis núcleo (si aún no está allí) para unirse a losLOSNeisseria elementos de respuesta hormonal enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ADN.
Interactúa con otros factores de transcripción para afectar la expresión génica (pueden actuar como potenciadores o inhibidores)
Vía de activación del receptor nuclear
Imagen por Kevin Ahern, editada por Lecturio.
Ejemplo de relevancia clínica
Síndrome de insensibilidad completa a losLOSNeisseria andrógenos: una afección recesiva ligada alALAmyloidosis cromosoma X enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la que una mutación genética afecta la función de losLOSNeisseria receptores de andrógenos, lo que lleva a la resistencia a la testosterona. LosLOSNeisseria receptores de andrógenos son receptores nucleares, localizados enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el citoplasma, que se mueven hacia el núcleo cuando se unen a losLOSNeisseria andrógenos y aumentan la transcripción de proteínas que causan efectos androgénicos. Con receptores anormales, losLOSNeisseria individuos tendrán un cariotipo 46,XY y testículos no descendidos, con genitales femeninos externos y desarrollo mamario (debido a la conversión periférica del exceso de testosterona enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum estrógeno).
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). Garland Science.
Nelson, D. L., & Cox, M. M. (2021). Lehninger principles of biochemistry (8th ed.). W. H. Freeman.
Rogers, J., et al. (2024). Mechanical control of antigen detection and discrimination by T and B cell receptors. Biophysical Journal, 123(15), 2234–2255. https://doi.org/10.1016/j.bpj.2024.05.020
Su, J., et al. (2024). Cell–cell communication: New insights and clinical implications. Signal Transduction and Targeted Therapy, 9(1), 1–52. https://doi.org/10.1038/s41392-024-01888-z
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Medical Premium le brinda acceso completo a todo el contenido y las funciones
Obtenga Premium para ver todos los vídeos
Verifica tu correo electrónico para obtener una prueba gratuita.
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Premium le ofrece acceso completo a todos los contenidos y funciones, incluido el banco de preguntas de Lecturio con preguntas actualizadas de tipo tablero.