Las funciones principales del tracto gastrointestinal incluyen la digestión de losLOSNeisseria alimentos y la absorción de nutrientes. Múltiples órganos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el sistema gastrointestinal secretan diversas sustancias hacia el lumen para ayudar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la digestión y/o regulación de la función gastrointestinal. La mayoría de las secreciones digestivas provienen de las glándulas salivales, estómago, páncreas y vesícula biliar, aunque losLOSNeisseria intestinos también secretan fluidos y moco, que son fundamentales para proteger sus paredes internas.
La digestión se divide enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum 3 fases, a saber, cefálica, gástrica e intestinal.
Fase cefálica
Comienza cuando el cerebro recibe aferencias sensoriales sobre la comida procedentes de losLOSNeisseria quimiorreceptores y mecanorreceptores situados enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las cavidades oral y nasal
Las aferencias sensoriales capaces de estimular la actividad gástrica incluyen:
Ver, oler, saborear o pensar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la comida
Masticar y tragar
Induce a losLOSNeisseria centros cerebrales superiores a estimular el complejo vagal dorsal
El nervio vago (parasimpático) libera acetilcolina, que conduce a:
↑ Secreciones salivales
↑ Secreciones gástricas (ácido clorhídrico (HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia), pepsinógeno) → digestión química
↑ Motilidad gástrica → digestión mecánica
Fase gástrica
Comienza cuando la comida ingerida entra alALAmyloidosis estómago
Estímulos químicos: e.g., la presencia de proteínas, péptidos y aminoácidos
Estímulos mecánicos: e.g., estiramiento
LosLOSNeisseria estímulos desencadenan varios reflejos → ↑ motilidad y secreciones gástricas
Reflejos cortos (reflejos locales):
Contenido dentro del sistema nervioso entérico
Las señales sensoriales viajan a losLOSNeisseria cuerpos celulares enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el sistema nervioso entérico (ubicado dentro de la pared intestinal).
El sistema nervioso entérico coordina la respuesta → envía una señal a través de losLOSNeisseria eferentes del sistema nervioso entérico→ la acetilcolina estimula ↑ secreciones gástricas y motilidad gastrointestinal
Reflejos largos (reflejos vago-vagales):
La respuesta se coordina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el cerebro.
Las señales sensoriales se transmiten a losLOSNeisseria aferentes del nervio vago → viajan alALAmyloidosis complejo vagal dorsal enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la médula
El complejo vagal dorsal coordina una respuesta → envía una señal a través de eferentes del nervio vago alALAmyloidosis estómago → la acetilcolina estimula las secreciones gástricas y la motilidad
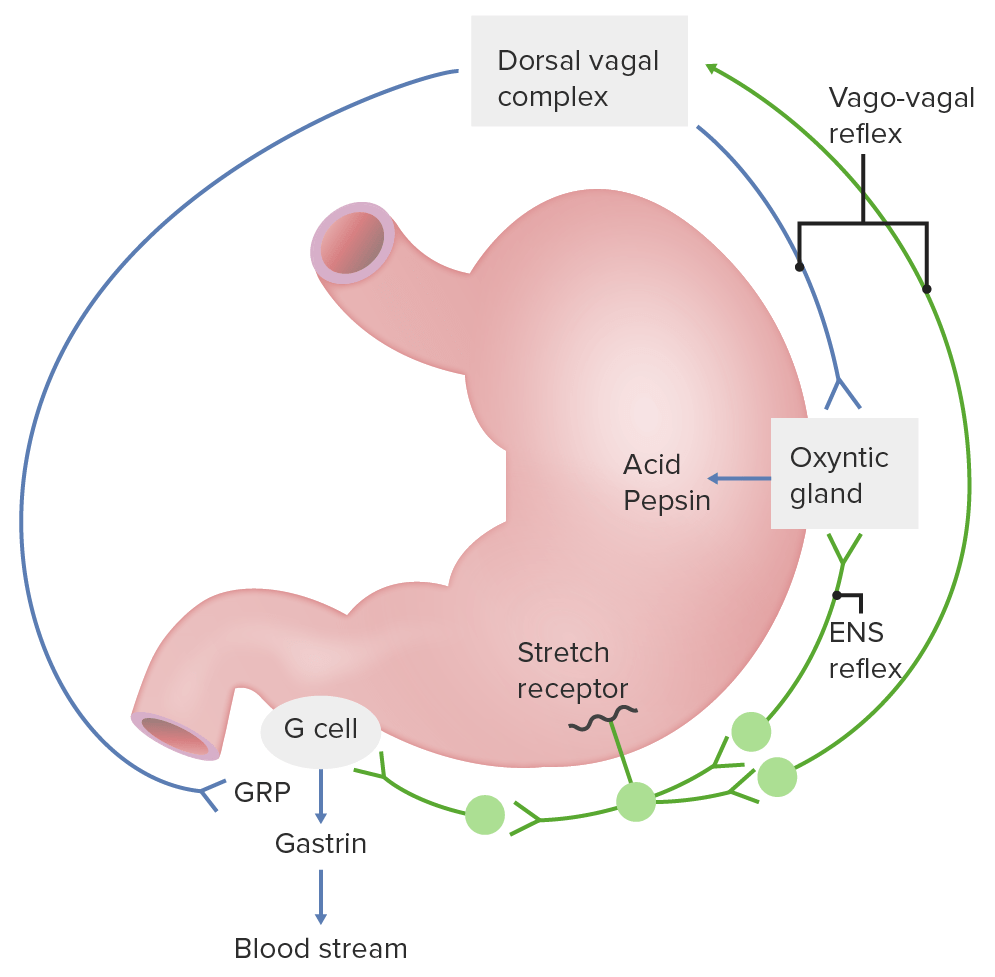
Regulación neural de las secreciones gástricas: Se muestran el reflejo del sistema nervioso entérico (ENS) (también conocido como reflejo corto) y el reflejo vasovagal (también conocido como reflejo largo). GRP: péptido liberador de gastrina
Imagen por Lecturio.
Fase intestinal
Comienza cuando la comida sale del estómago y entra alALAmyloidosis duodeno
Duodeno:
Modula la actividad gástrica a través de hormonas y reflejos neurales:
Inicialmente, las señales estimulan la actividad gástrica.
Pronto, las señales inhiben la actividad gástrica.
Secreta moléculas de señalización (e.g., colecistoquinina) que estimulan las secreciones del páncreas y la vesícula biliar
Comienza la absorción de nutrientes.
Fases de la digestión con sus componentes funcionales
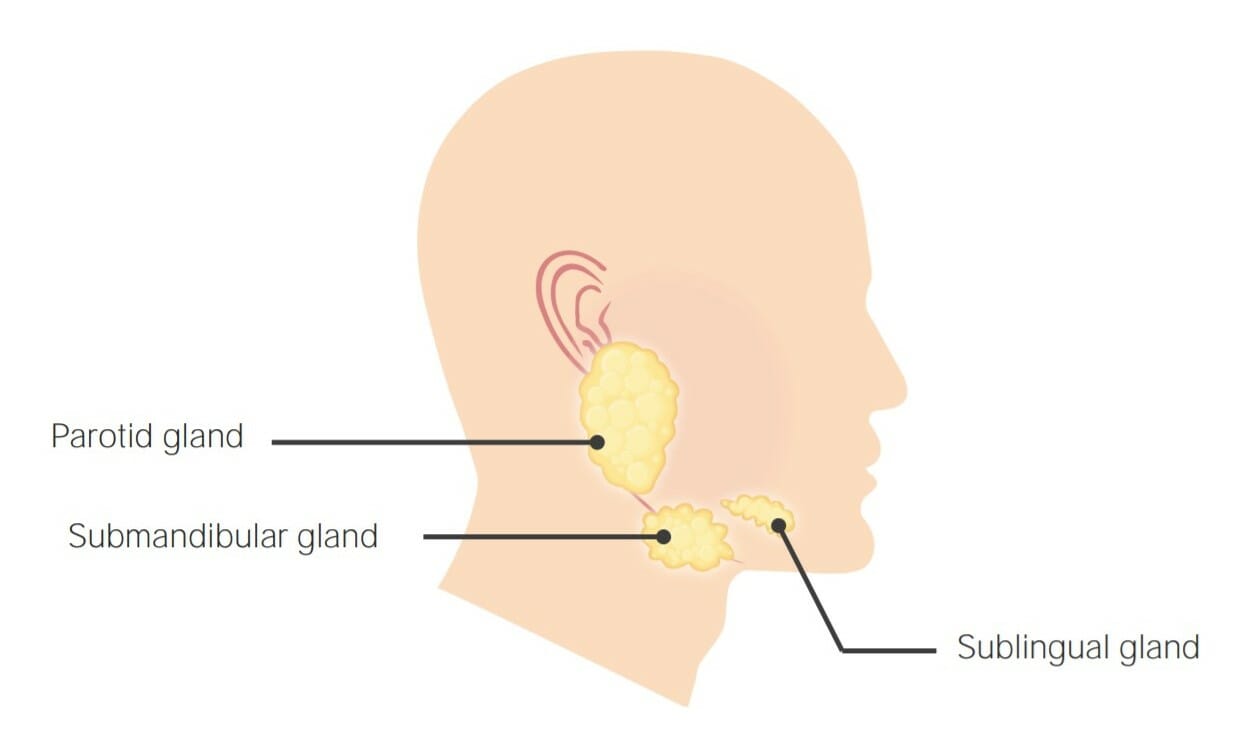
Hay 3 glándulas salivales primarias (todas con una estructura tubuloacinar), que juntas producen una combinación de secreciones serosas y mucosas.
Glándulas parótidas: secreción serosa
Glándulas submandibulares: secreciones serosas y mucosas
Glándulas sublinguales: principalmente secreciones mucosas, con un pequeño componente seroso
Localización de las 3 glándulas salivales primarias: Glándulas parótidas, submaxilares y sublinguales
Imagen por Lecturio.
Constituyentes de la salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy
La salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy se compone de:
Agua
Moco
Electrolitos:
K+: ayuda enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la reabsorción de Na+ y agua
HCO3–: amortiguador de ácidos
Enzimas:
Amilasa salival: digestión del almidón
Lipasa lingual: digestión de lípidos
Agentes antimicrobianos:
Muramidasa: lisozima (una enzima que puede destruir las paredes celulares de ciertas bacterias)
Lactoferrina: se une alALAmyloidosis hierro, lo que ayuda a prevenir el crecimiento bacteriano
Inmunoglobulina A (IgAIgARepresents 15-20% of the human serum immunoglobulins, mostly as the 4-chain polymer in humans or dimer in other mammals. Secretory iga is the main immunoglobulin in secretions.Immunoglobulins: Types and Functions): mediador inmunitario (anticuerpo secretado)
Producción de salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy
Descripción general:
La salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy es producida por células acinares enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las glándulas salivales como un filtrado.
Modificada por células ductales a medida que se mueve a través de losLOSNeisseria conductos
Las glándulas salivales secretan aproximadamente de 1‒1,5 L de salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: AnatomyalALAmyloidosis día.
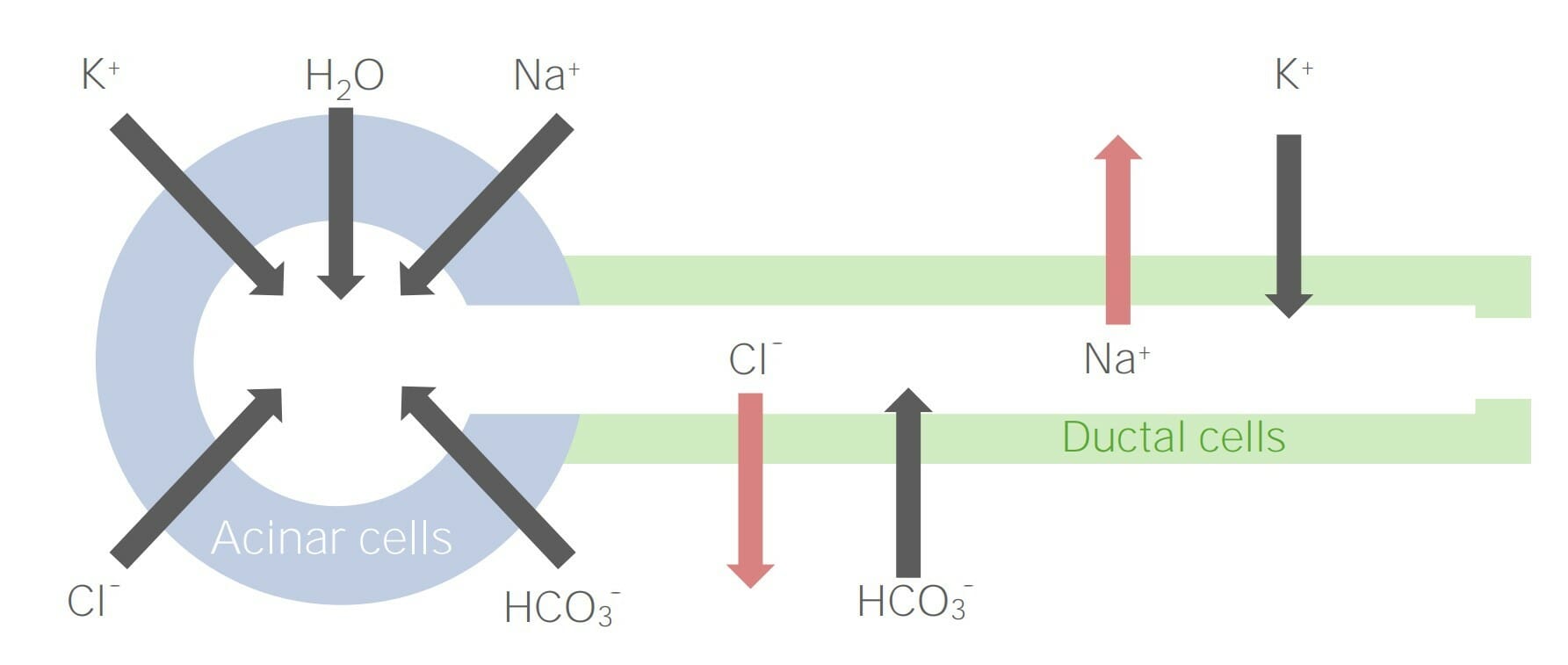
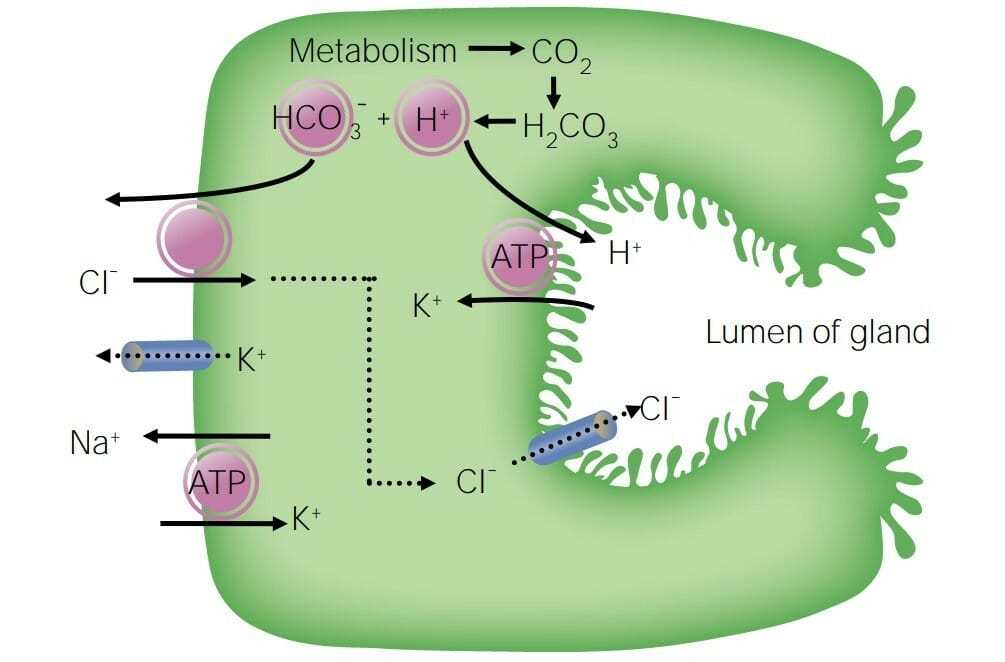
Diagrama que detalla la secreción iónica de las células acinares y su movimiento a través de las células ductales
Imagen por Lecturio.
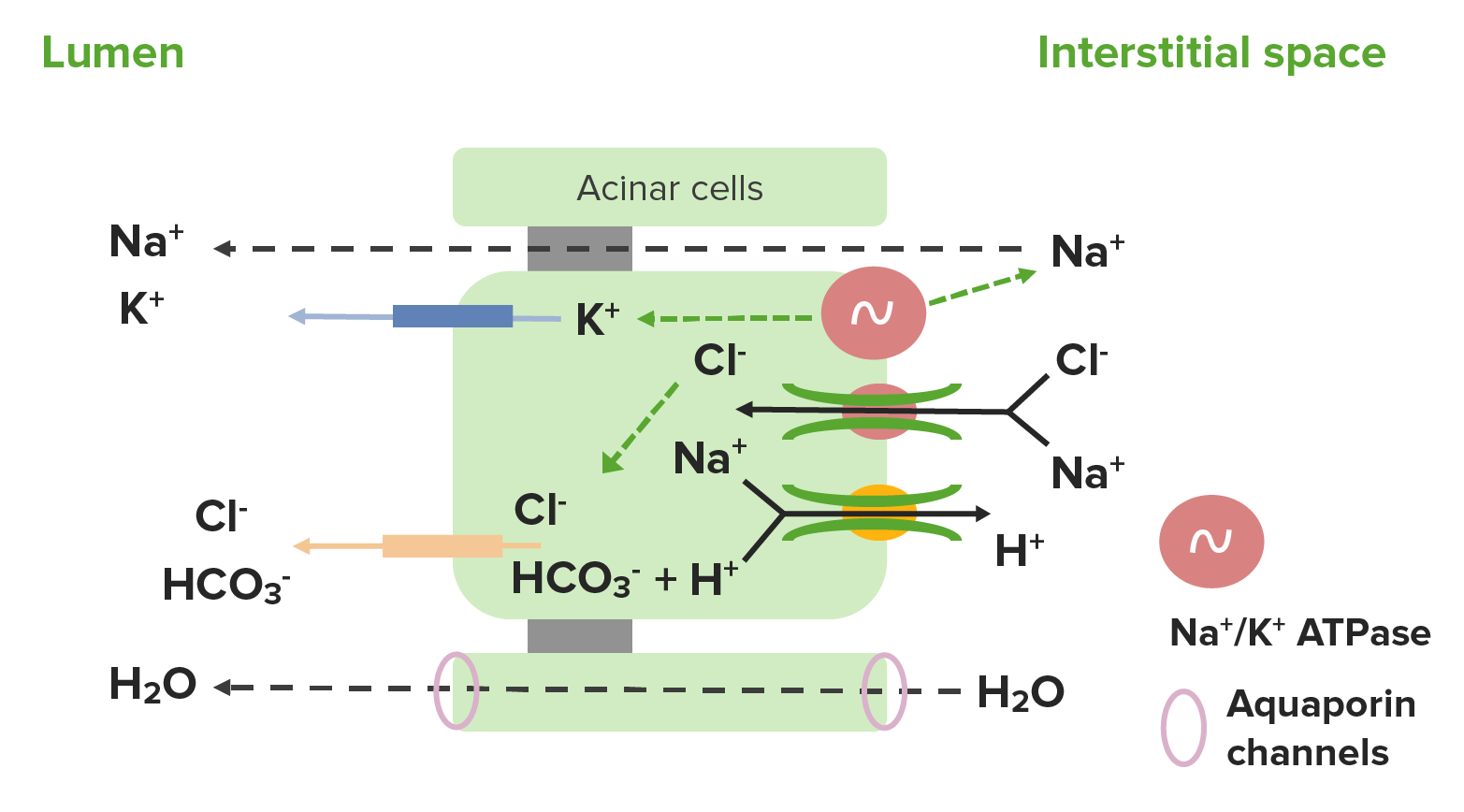
Células acinares:
Las células acinares secretan un filtrado que contiene Na+, K+, Cl–, HCO3–, agua y otras sustancias.
La bomba de Na+/K+ ATPasa mantiene:
K+ concentrado dentro de las células acinares
Na+ concentrado enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el espacio intersticial
Movimiento de Na+:
El Na+ se mueve paracelularmente hacia el lumen de losLOSNeisseria acinos a favor de su gradiente de concentración → el Na+ se secreta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy
Se introduce enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células acinares a través del cotransportador Na+/Cl–enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana basolateral → el Na+ se transporta alALAmyloidosis espacio intersticial con el H+
Movimiento de agua:
El agua sigue alALAmyloidosis Na+ hacia el lumen a través de losLOSNeisseria canales de acuaporina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células acinares.
Movimiento isosmótico
Movimiento de K+:
Concentrado dentro de las células acinares debido a la acción de la bomba Na+/K+ ATPasa
Se mueve a través de losLOSNeisseria canales de K+ por su gradiente de concentración hacia el lumen
Movimiento de Cl–, H+, y HCO3–:
El CO2 se produce durante el metabolismo → CO2 se combina con H2O → ácido carbónico (H2CO3) → se divide enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum H+ y HCO3–
El H+ y el Na+ se transportan alALAmyloidosis espacio intersticial a través del cotransportador H+/Na+enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana basolateral.
El HCO3– (del metabolismo) y el Cl– (del cotransportador basolateral Na+/Cl–) se secretan a través del cotransportador HCO3–/Cl–enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana apical (luminal)
Producen y secretan otras sustancias (e.g., mucina, enzimas) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria ácinos
Diagrama que muestra la secreción de iones por las células acinares: En las células acinares, el Na+, el K+, el Cl– y el HCO3– se filtran o secretan en el líquido salival (nota: el Na+ y el Cl– son luego reabsorbidos por las células ductales).
Imagen por Lecturio.
Células ductales:
Las células ductales modifican el filtrado a medida que se mueve por medio de losLOSNeisseria conductos, reabsorbiendo finalmente Na+ y Cl– y secretando más K+ y HCO3–.
Reabsorción de Na+ y secreción de K+:
El Na+ y el H+ se reabsorben a través de la membrana apical por medio del cotransportador Na+/H+.
El Na+ se bombea a través de la membrana basolateral hacia el espacio intersticial por medio de la bomba Na+/K+ ATPasa.
El H+ se recicla de vuelta alALAmyloidosis lumen, mientras que el K+ se secreta: el H+ y el K+ se mueven a través de la membrana apical por medio del cotransportador H+/K+.
Reabsorción de Cl– y secreción de HCO3–:
El CO2 se produce durante el metabolismo → CO2 se combina con H2O → H2CO3 → se divide enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum H+ y HCO3–
El HCO3– se intercambia por Cl–enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana apical (el HCO3– se secreta y el Cl– se reabsorbe enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la célula ductal) a través del contratransportador HCO3–/Cl–.
El Cl– se reabsorbe enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el espacio intersticial a través de losLOSNeisseria canales de Cl–.
El H+ (residuo de la reacción de la anhidrasa carbónica) se elimina a través de la membrana basolateral a través del contratransportador H+/Na+ (el Na+ desciende por su gradiente de concentración hacia la célula).
Como el agua no se reabsorbe (pero algunos iones sí), la salivaSalivaThe clear, viscous fluid secreted by the salivary glands and mucous glands of the mouth. It contains mucins, water, organic salts, and ptyalin.Salivary Glands: Anatomy resultante es hipotónica.
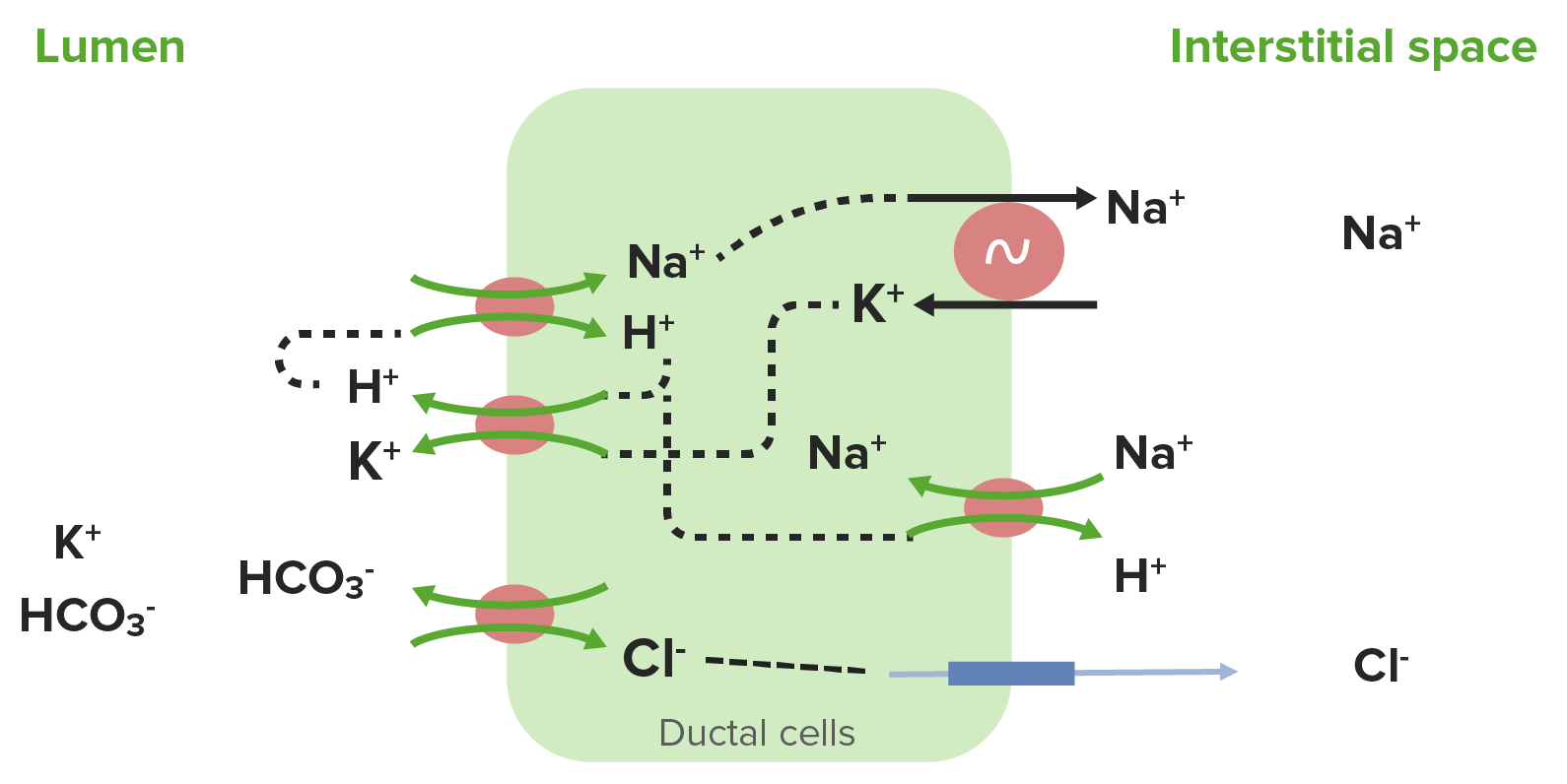
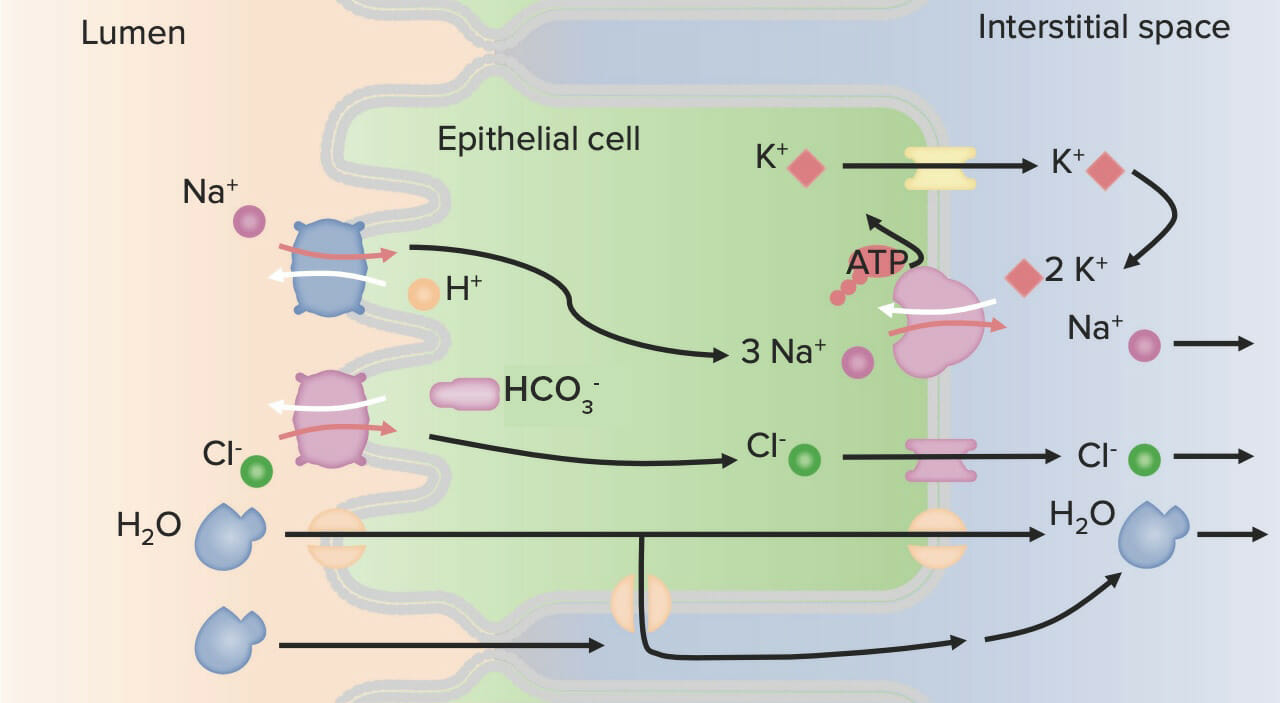
Diagrama que muestra el transporte de iones por células ductales: El cotransportador Na+/H+ reabsorbe Na+ e H+ del fluido salival. Luego, el Na+ es bombeado a través de la membrana basolateral por la Na+/K+ ATPasa, y el K+ entra en la célula. A continuación, el H+ se recicla de vuelta hacia el lumen junto con el K+ a través de un cotransportador H+/K+. Luego, el H+ se usa para reabsorber más Na+, mientras que el K+ permanece en la saliva y se excreta. El cloruro se reabsorbe, mientras que el HCO3– se excreta a través del intercambiador Cl–/HCO3– en la membrana apical.
Imagen por Lecturio.
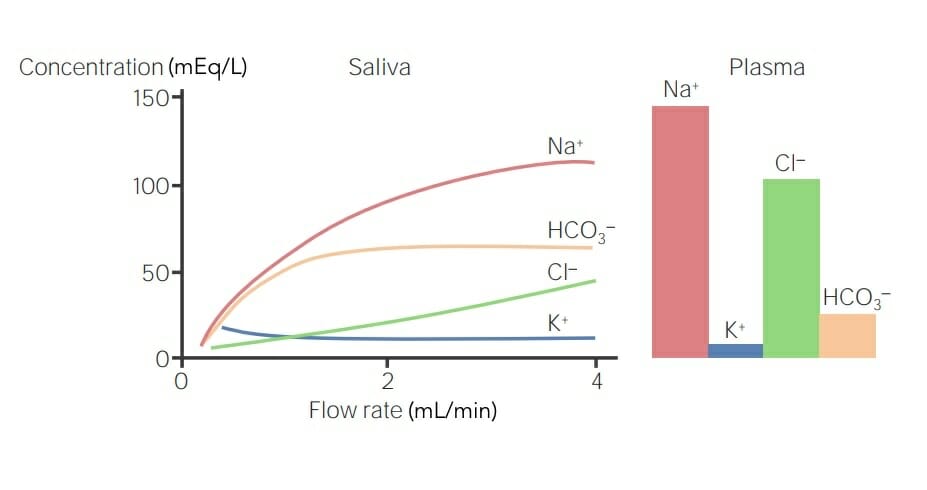
Secreción salival de iones y sus niveles plasmáticos: Si la concentración del ion en el plasma es mayor que la de la saliva, el ion se reabsorbe. Si la concentración del ion en el plasma es menor que la de la saliva, el ion se secreta. Cuanto más rápido el flujo de saliva, menos tiempo hay para la secreción o reabsorción de iones, lo que afecta sus concentraciones salivales. Concentraciones más altas de Na+ y Cl– están presentes en el plasma que en la saliva y, por lo tanto, se reabsorben, mientras que el K+ y HCO3– están presentes en concentraciones más bajas en el plasma que en la saliva y, por lo tanto, se secretan.
Imagen por Lecturio.
Control y regulación de las secreciones salivales
Participa enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase cefálica de la digestión:
Aferencias sensoriales capaces de estimular la salivación:
Pensar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la comida
Ver, oler o probar la comida
Induce a losLOSNeisseria centros cerebrales superiores a estimular el sistema nervioso autónomo (SNA) (principalmente losLOSNeisseria parasimpáticos)
La estimulación parasimpática aumenta las secreciones salivales a través de:
Acetilcolina: aumenta las secreciones salivales
Péptido intestinal vasoactivo: aumenta el flujo sanguíneo a las glándulas salivales
Estimulación simpática: aumenta las secreciones salivales enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum menor grado a través de la liberación de norepinefrina
Relevancia clínica de las secreciones salivales
Xerostomía, o boca seca, es el término clínico utilizado para identificar la secreción salival alterada, que ocurre comúnmente como parte del síndrome de Sjögren, como un efecto secundario de algunos medicamentos (como antidepresivos, antihipertensivos o anticolinérgicos) y enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum personas que reciben radioterapia para losLOSNeisseria cánceres de cabeza y cuello.
Proteger el resto del sistema gastrointestinal matando a la mayoría de losLOSNeisseria microbios
Preparación del quimo para el intestino delgado a través de:
Desintegración mecánica
Digestión química por ácido y pepsinógeno
Absorción de sustancias lipofílicas
Almacenamiento y liberación gradual de material enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el duodeno (regula la entrada de alimentos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el intestino delgado)
Glándulas oxínticas gástricas: células y sus secreciones
Las glándulas oxínticas gástricas se encuentran debajo (y desembocando enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum) las fosas gástricas. Las glándulas contienen numerosos tipos de células, que incluyen:
Células mucosas superficiales:
Recubren las fosas gástricas
Secretan bicarbonato y moco insoluble:
Forman una barrera protectora contra el ambiente ácido del estómago
Concentran bicarbonato enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el moco
Células mucosas del cuello:
Situadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el cuello de las glándulas donde se unen a las fosas gástricas
Secretan moco soluble
Células madre:
Se encuentran entre las fosas y la entrada de las glándulas
Producen nuevas células para reemplazar tanto las células mucosas superficiales enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las fosas como las células glandulares debajo
Las células epiteliales del estómago se reemplazan cada 3‒6 días.
Células parietales:
Situadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la región media-inferior de las glándulas
Secretan:
HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia
Factor intrínseco: importante para la absorción de vitamina B12
Células principales:
Células glandulares más numerosas
Situadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la región media-inferior de las glándulas
Secretan:
Pepsinógeno → convertido a su forma activa pepsina por el HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia→ descompone las proteínas
Lipasa gástrica → descompone las grasas
Células enteroendocrinas:
Situadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la base de las glándulas
Las células D secretan somatostatina:
Inhibe muchas secreciones.
Liberada enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum respuesta a H+ (la forma natural de “apagar” la producción de ácido)
Las células G secretan gastrina:
Estimula a las células parietales para que secreten HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia
Tiene efectos tróficos/de crecimiento enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la mucosa gastrointestinal
Liberada enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum respuesta a proteínas, péptidos y aminoácidos
Las células tipo enterocromafines secretan histamina (que estimula a las células parietales a secretar HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia).
Otros mensajeros químicos secretados por las células enteroendocrinas:
Sustancia P
Péptido intestinal vasoactivo
Secretina
Neuropéptido Y
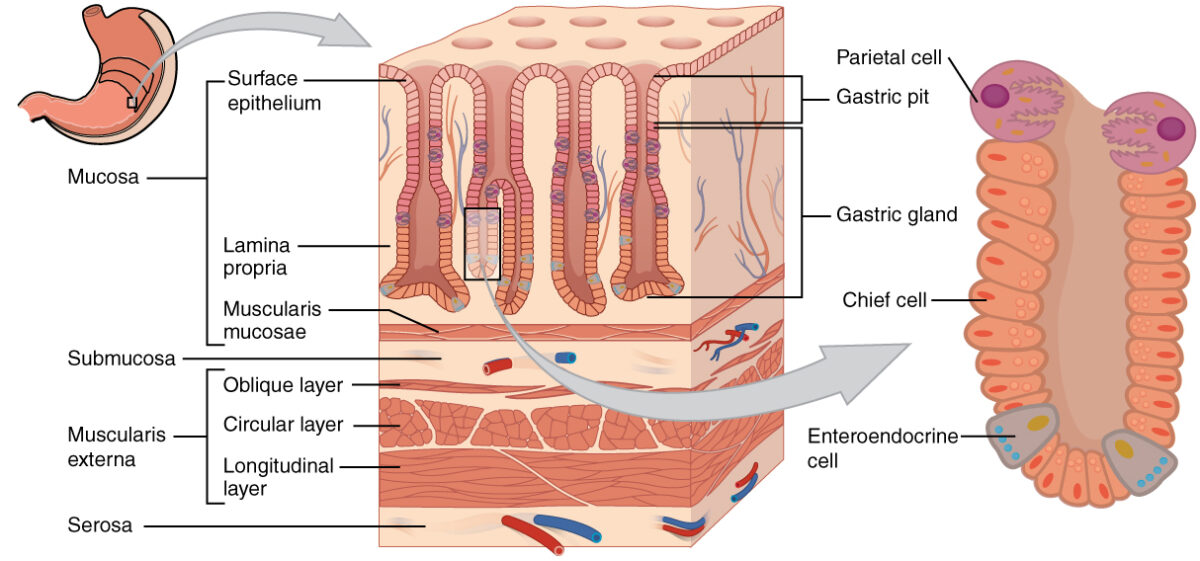
Capas de la pared del estómago: En el epitelio, las fosas gástricas conducen a glándulas gástricas que secretan una variedad de sustancias para ayudar en la digestión.
Imagen: “The stomach wall is adapted for the functions of the stomach”por OpenStax College. Licencia: CC BY 4.0
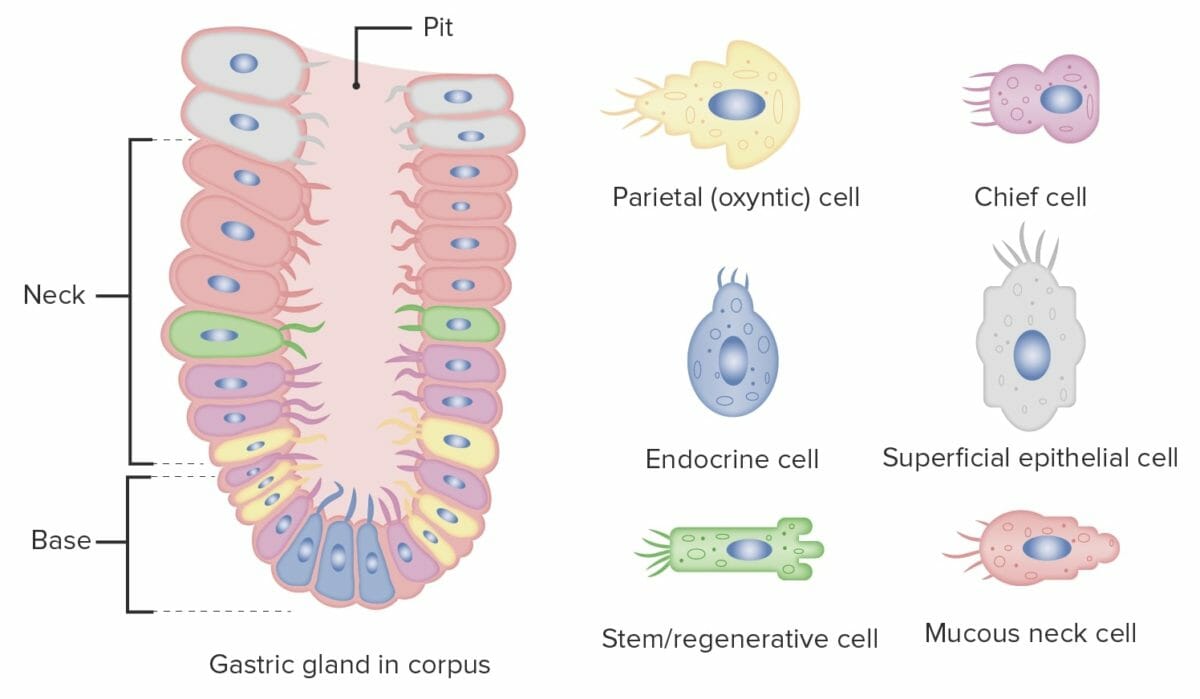
Estructura de una glándula gástrica con sus diferentes tipos de células
Imagen por Lecturio.
Producción y secreción de ácido enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células parietales
El metabolismo normal produce CO2 → se combina con H2O → H2CO3 → se divide enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum H+ + HCO3–
H+ es bombeado hacia el lumen a cambio de K+ por la H+/K+ ATPasa:
Se introduce 1 molécula de K+enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la célula.
El K+ sale de la célula a través de la membrana basolateral siguiendo su gradiente de concentración a través de un canal de K+.
HCO3– se intercambia con Cl– a través de la membrana basolateral:
HCO3– se mueve hacia el espacio intersticial.
Cl– se mueve hacia la célula → se mueve hacia el lumen a través de su propio canal
Resultado final:
H+ y Cl– se secretan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el lumen.
K+ y HCO3– se mueven alALAmyloidosis espacio intersticial.
Movimiento de iones en las células parietales: El ácido carbónico se disocia en H+ y HCO3–. El H+ es intercambiado por K+ en la membrana apical por la H+/K+ ATPasa. Un HCO3– se intercambia por Cl– en la membrana basolateral; el Cl– luego se mueve hacia el lumen.
Imagen por Lecturio.
Control y regulación de las secreciones gástricas
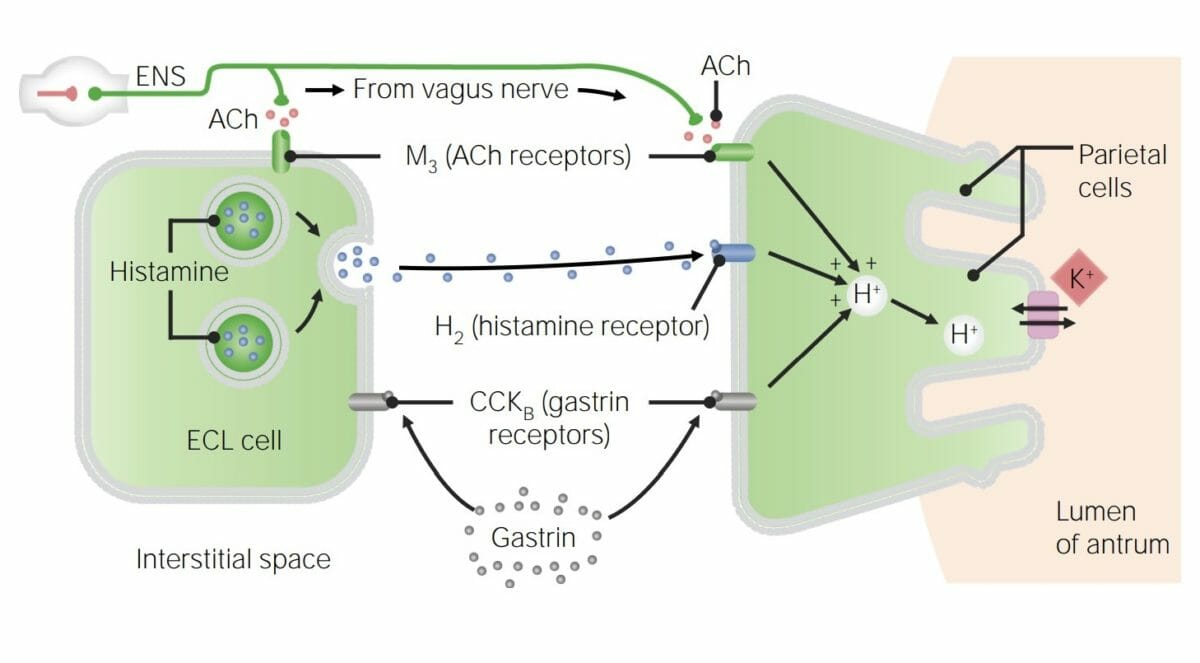
Las secreciones gástricas están muy influenciadas por la señalización parasimpática a través del nervio vago (nervio craneal X), que libera acetilcolina que induce la producción de ácido por varias vías.
Producción de secreciones:
40% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase cefálica
50% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase gástrica
10% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase intestinal
La secreción de HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia de las células parietales es estimulada por:
Acetilcolina de losLOSNeisseria nervios vago y mientérico (vías reflejas largas y cortas, respectivamente)
Gastrina de las células G enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el estómago
Histamina de las células tipo enterocromafines enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el estómago
La secreción de HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia es inhibida por:
Somatostatina
Prostaglandinas
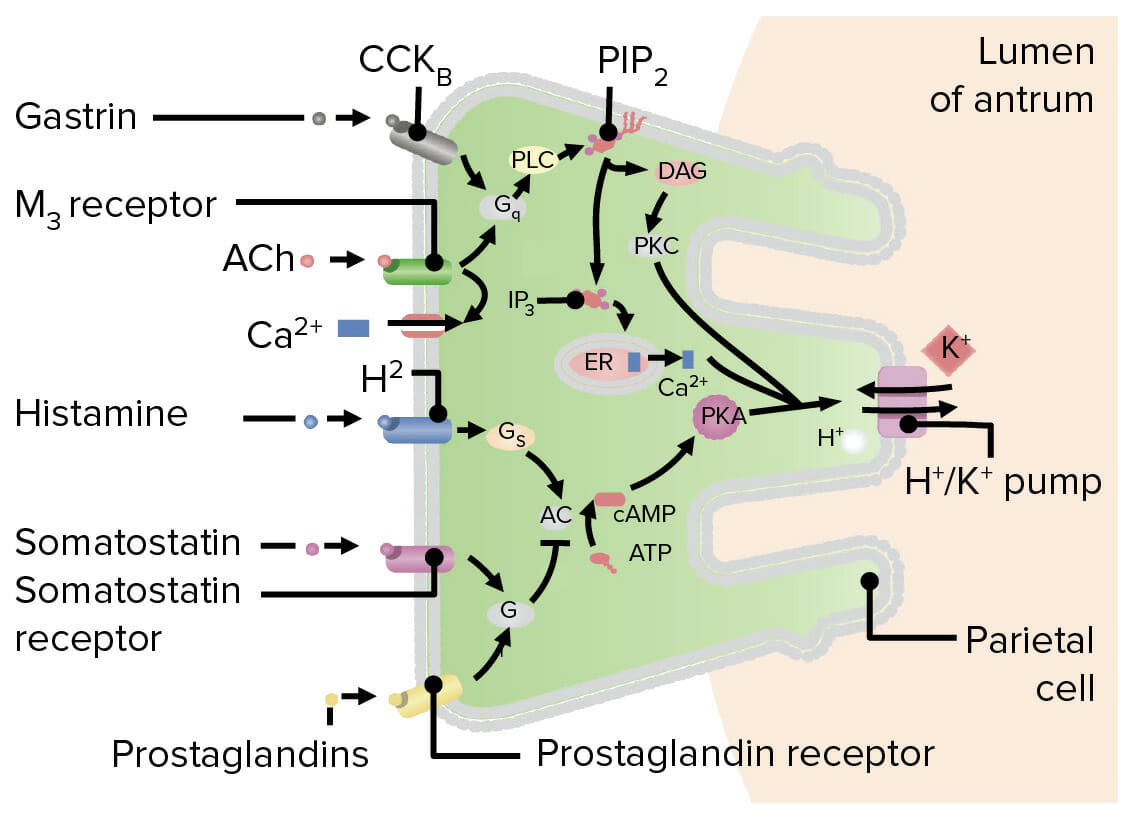
Vías de estimulación de ácido
Víadirecta (activación directa de las células parietales):
La acetilcolina estimula losLOSNeisseria receptores muscarínicos (M3) de las células parietales.
Activa Gq (una proteína G)
Gq activa la fosfolipasa C.
La fosfolipasa C escinde el fosfatidilinositol-4,5-bisfosfato (PIP2) para producir:
Trifosfato de inositol (IP3) → liberación de calcio (CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+) del retículo endoplásmico
Diacilglicerol → fosforilación de proteína quinasa C
Tanto el CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ como la proteína quinasa C activan la H+/K+ ATPasa para secretar H+.
Vía de la gastrina:
La acetilcolina estimula a las células G para que liberen gastrina.
La gastrina activa losLOSNeisseria receptores de colecistoquinina B enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células parietales.
La colecistoquinina B activa la fosfolipasa C→ escinde el fosfatidilinositol-4,5-bisfosfato (PIP2) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum trifosfato de inositol (IP3) + diacilglicerol → ↑ CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ + proteína quinasa C → ↑ actividad de la H+/K+ ATPasa
Vía de la histamina:
La acetilcolina estimula las células tipo enterocromafines para que liberen histamina.
La histamina activa losLOSNeisseria receptores H2enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células parietales.
La adenilato ciclasa convierte adenosin trifosfato (ATP) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum adenosin monofosfato ciclico (AMPc).
El AMPc fosforila/activa la proteína quinasa A.
La proteína quinasa A estimula la H+/K+ ATPasa para secretar H+.
LosLOSNeisseria efectos de estas vías son de naturaleza potenciadora/sinérgica → La actividad de acetilcolina + gastrina + histamina simultáneamente da como resultado una mayor secreción de HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia que la suma de la secreción de HClHCLHairy cell leukemia (HCL) is a rare, chronic, B-cell leukemia characterized by the accumulation of small mature B lymphocytes that have “hair-like projections” visible on microscopy. The abnormal cells accumulate in the peripheral blood, bone marrow (causing fibrosis), and red pulp of the spleen, leading to cytopenias.Hairy Cell Leukemia si cada una actuara sola
Diagrama que muestra las vías de estimulación e inhibición de la secreción de ácido en una célula parietal: La gastrina y la acetilcolina activan Gq, una proteína G, que escinde el fosfatidilinositol-4,5-bisfosfato (PIP2) en trifosfato de inositol (IP3) y diacilglicerol (DAG). El trifosfato de inositol provoca la liberación de calcio (Ca2+) del retículo endoplásmico, mientras que el diacilglicerol activa la proteína quinasa C (PKC) a través de la fosforilación. Tanto el Ca2+ como la proteína quinasa C estimulan la bomba H+/K+ para secretar ácido. La histamina activa Gs, que activa la adenilato ciclasa (AC), lo que da como resultado un aumento del AMPc intracelular. A continuación, el AMPc activa la proteína cinasa A (PKA), que estimula la bomba H+/K+ para secretar ácido. La somatostatina y las prostaglandinas inhiben la producción de ácido al inducir Gi, que inhibe la adenilato ciclasa. CCK: colecistoquinina PLC: fosfolipasa C Ach: acetilcolina
Imagen por Lecturio.
Diagrama que detalla las vías directas e indirectas de estimulación de la liberación de ácido por la acetilcolina (ACh): La vía directa implica la estimulación de los receptores muscarínicos (M3) en las células parietales. Las vías indirectas implican la estimulación de células tipo enterocromafines (ECL), que inducen la liberación de histamina, y la estimulación de células G que las inducen a secretar gastrina. Tanto la histamina como la gastrina estimulan las células parietales para que secreten ácido. ENS: sistema nervioso entérico CCK: colecistoquinina
Imagen por Lecturio.
Vías de inhibición del ácido
Somatostatina y prostaglandinas:
La somatostatina (liberada de las células D enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el estómago) y las prostaglandinas activan la proteína inhibidora Gi.
Gi inhibe la adenilato ciclasa → losLOSNeisseria niveles de AMPc caen → ↓ activación de la proteína quinasa A → ↓ actividad H+/K+ ATPasa
Relevancia clínica de las secreciones gástricas
Enfermedad por reflujo gastroesofágico (ERGE): ocurre cuando el ácido del estómago regresa con frecuencia hacia el esófago. El retrolavado (reflujo ácido) puede irritar el revestimiento del esófago, lo que provoca síntomas como ardor retroesternal (acidez estomacal) y, con el tiempo, puede provocar inflamación (esofagitis), metaplasiaMetaplasiaA condition in which there is a change of one adult cell type to another similar adult cell type.Cellular Adaptation (esófago de Barrett) y progresión a cáncer de esófago. La ERGE sin complicaciones se puede controlar con cambios enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el estilo de vida y medicamentos de venta libre.
Esófago de Barrett: una afección caracterizada por cambios metaplásicos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el epitelio escamoso estratificado normal del esófago a epitelio cilíndrico. El cambio es una consecuencia de la ERGE crónica y se considera premaligno.
Medicamentos para reducirlas secreciones deácido gástrico:losLOSNeisseria medicamentos incluyen inhibidores de la bomba de protones (IBP) y antagonistas de losLOSNeisseria receptores H2 y están indicados con mayor frecuencia para tratar la enfermedad de úlcera péptica, la ERGE y la dispepsia. El mecanismo de acción de losLOSNeisseria IBP para reducir el ácido gástrico es mediante la inhibición de la H+/K+ ATPasa enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las células parietales, mientras que el de losLOSNeisseria bloqueadores H2 es mediante la inhibición de losLOSNeisseria efectos estimulantes de la histamina sobre las células parietales.
Síndrome de Zollinger-Ellison: un tumorTumorInflammation secretor de gastrina (a menudo maligno) que surge del páncreas, estómago, duodeno, yeyuno y/o ganglios linfáticos, que se caracteriza por úlceras pépticas recurrentes/refractarias, reflujo gastroesofágico y diarrea. El diagnóstico se basa enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum niveles elevados de gastrina sérica enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum ayunas. El tratamiento es con resección quirúrgica del tumorTumorInflammation y/o tratamiento sintomático.
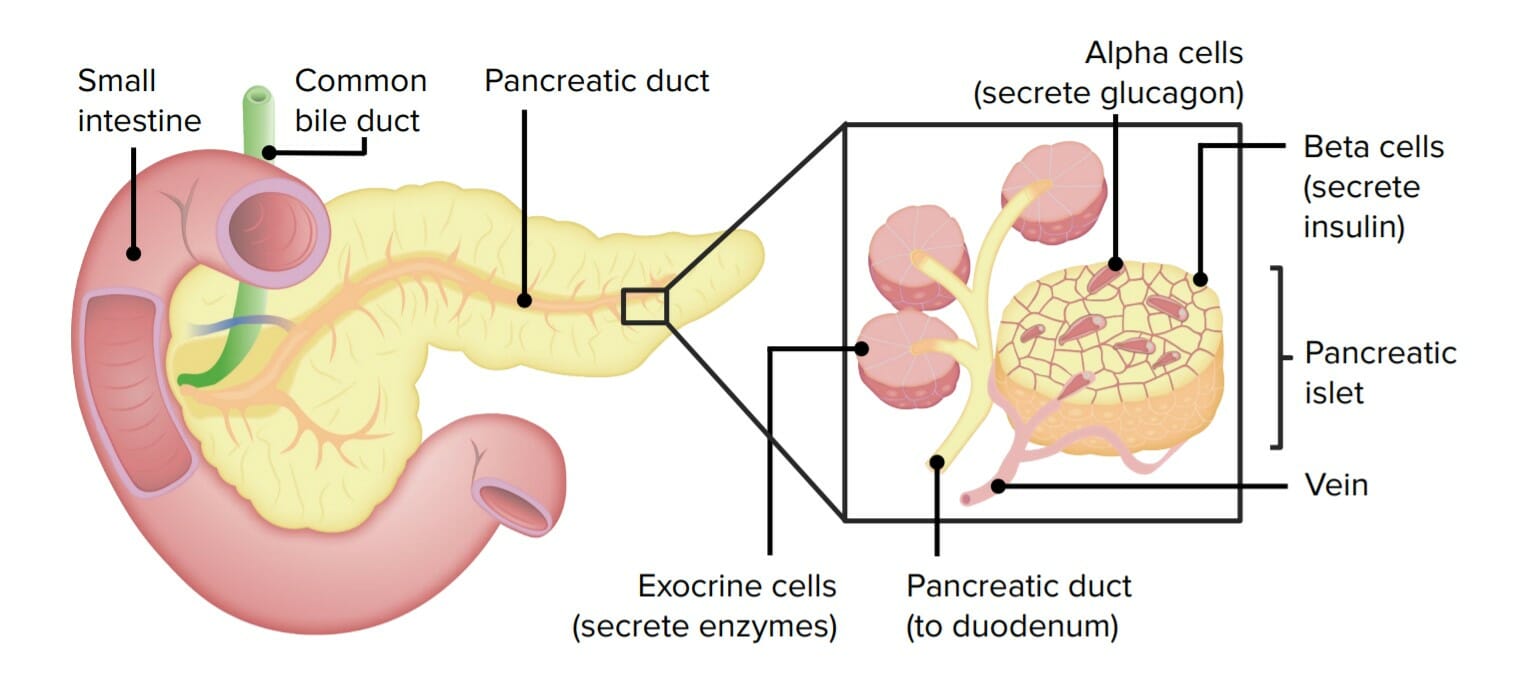
Libera enzimas pancreáticas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el duodeno
Dispuesto como grupos de acinos que drenan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum un sistema ductal → conductos pancreáticos principal y accesorio → duodeno
Endocrinos:
Libera hormonas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el torrente sanguíneo
Las células están ubicadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum grupos conocidos como islotes.
Células α: secreción de glucagón
Células β: secreción de insulina
Células 𝛿: secreción de somatostatina
Un dibujo del páncreas que identifica sus 2 componentes tisulares principales: el páncreas endocrino (islotes de Langerhans o islotes pancreáticos) y el páncreas exocrino (células exocrinas o acinos pancreáticos)
Imagen por Lecturio.
Secreciones del páncreas exocrino
El páncreas exocrino secreta una mezcla conocida como jugo pancreático, que contiene agua, enzimas, zimógenos (proteínas inactivas), HCO3– y electrolitos:
Tampón (neutraliza el ácido del estómago): HCO3–
Para la digestión de carbohidratos: amilasa pancreática
Para la digestión de lípidos:
Lipasa pancreática
Fosfolipasa A2
Colesterol esterasa
Para la digestión de proteínas y péptidos (secretados principalmente como zimógenos):
Tripsinógeno → activado por la enteropeptidasa (anteriormente enteroquinasa) a tripsina
Quimotripsinógeno → activado por tripsina a quimotripsina
Procarboxipeptidasa → activada por tripsina a carboxipeptidasa
Proelastasa → activada por tripsina elastasa
Para la digestión de nucleótidos:
Ribonucleasa (RNAsa)
Desoxirribonucleasa (ADNasa)
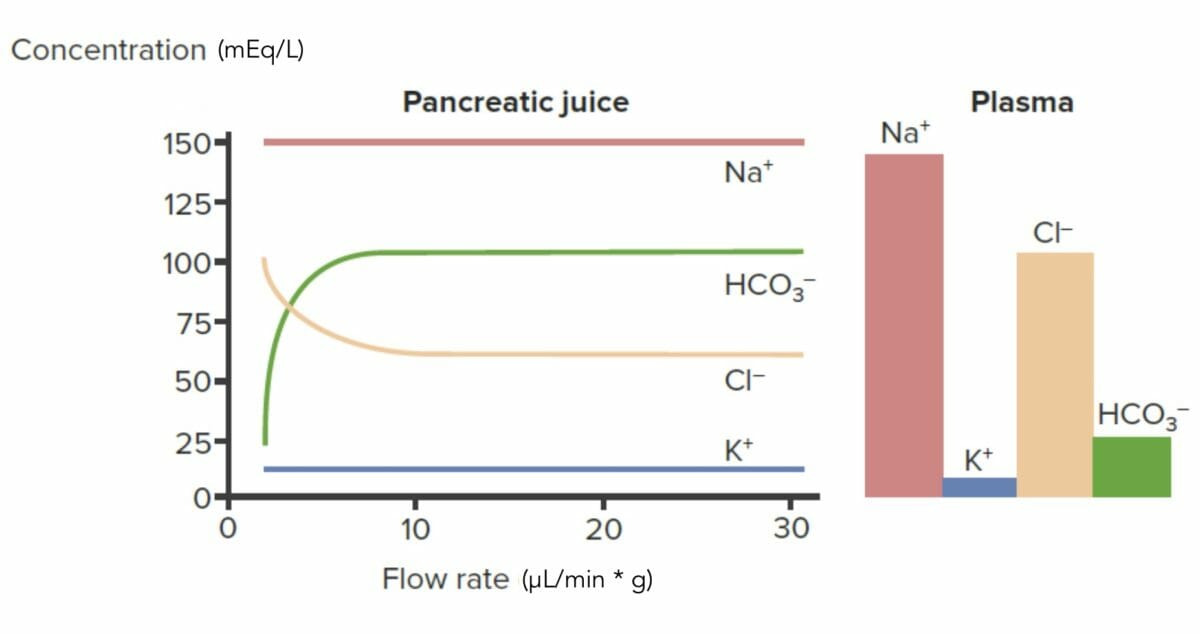
Concentraciones de iones enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el jugo pancreático
HCO3–:
Secretado activamente: concentración enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum jugo pancreático > que enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumplasmaPlasmaThe residual portion of blood that is left after removal of blood cells by centrifugation without prior blood coagulation.Transfusion Products
La secreción aumenta a medida que aumenta la tasa de flujo.
Cl–:
Reabsorbido activamente: concentración enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum jugo pancreático < que enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumplasmaPlasmaThe residual portion of blood that is left after removal of blood cells by centrifugation without prior blood coagulation.Transfusion Products
La reabsorción cae a medida que aumenta la tasa de flujo.
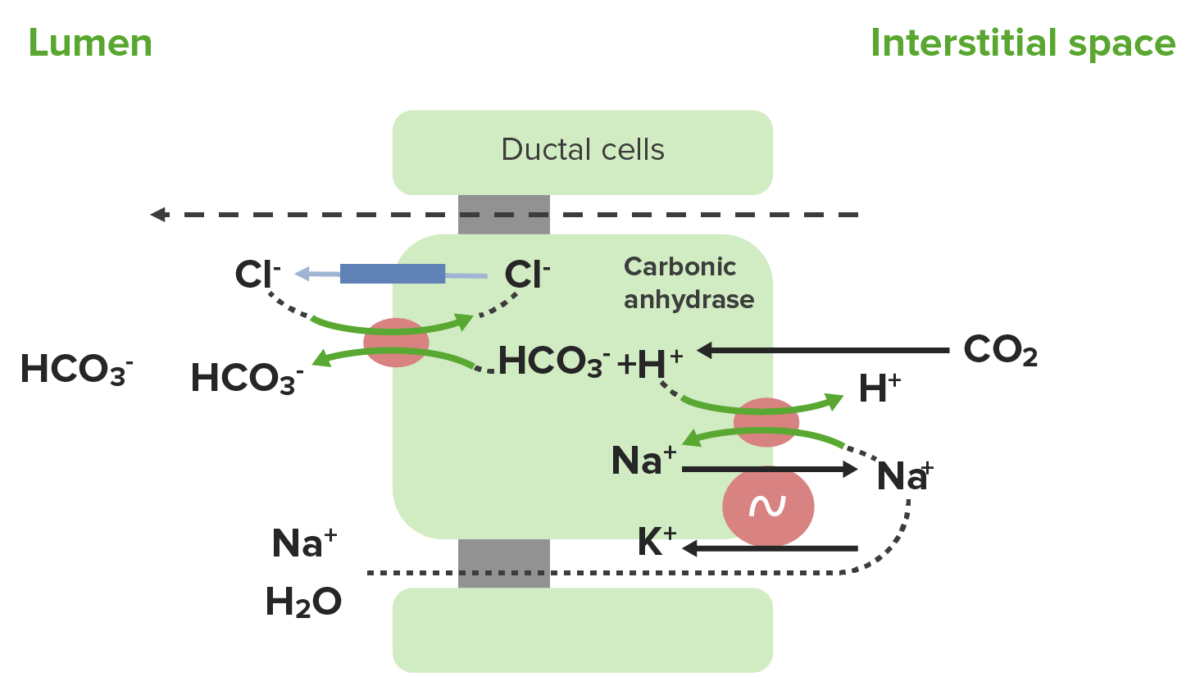
Mecanismo de secreción de HCO3– y reabsorción de Cl–:
El CO2 ingresa a las células → se combina con el agua para formar H2CO3 → se divide enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum H+ y HCO3–
El H+ se mueve de vuelta a través de la membrana basolateral hacia el espacio intersticial a través del intercambiador H+/Na+.
El HCO3– se secreta a través de la membrana apical hacia el lumen a través del intercambiador HCO3–/Cl–.
El Cl– se puede reciclar de vuelta alALAmyloidosis lumen a través del canal de Cl–.
El Na+ se elimina de la célula a través de la membrana basolateral a través del intercambiador Na+/K+ ATPasa.
El Na+ y el K+ no se secretan ni se reabsorben activamente:
La concentración enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el jugo pancreático es similar a la del plasmaPlasmaThe residual portion of blood that is left after removal of blood cells by centrifugation without prior blood coagulation.Transfusion Products.
Algo de Na+ se mueve paracelularmente hacia el lumen.
Secreción pancreática de iones y sus niveles plasmáticos: Obsérvese que a medida que aumenta la tasa de flujo (eje x), la concentración de HCO3– en la secreción pancreática (eje y) aumenta por encima de los niveles plasmáticos, lo que indica secreción. Sucede lo contrario con el Cl–, donde su concentración cae por debajo de los niveles plasmáticos, lo que indica reabsorción.
Imagen por Lecturio.
Secreción de HCO3– por las células ductales pancreáticas: el CO2 ingresa a las células, se combina con agua para formar ácido carbónico (H2CO3) y luego se divide en H+ y HCO3–. El H+ se mueve de vuelta a través de la membrana basolateral hacia el espacio intersticial a través de un intercambiador H+/Na+, mientras que el HCO3– se secreta a través de la membrana apical hacia el lumen a través de un intercambiador HCO3–/Cl–. El Cl– se puede reciclar de vuelta al lumen a través de un canal de Cl–. A continuación, el Na+ se elimina de la célula a través de la membrana basolateral a través de la Na+/K+ ATPasa. Parte del Na+ se desplaza paracelularmente hacia el lumen, trayendo consigo agua.
Imagen por Lecturio.
Control y regulación
Porcentaje de secreciones producidas:
25% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase cefálica, estimulada principalmente por la acetilcolina liberada por el nervio vago
10% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase gástrica, estimulada principalmente por reflejos vago-vagales
65% enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la fase intestinal, estimulada por la secretina y la colecistoquinina, ambas hormonas liberadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el duodeno
Estimulación de secreciones:
Liberación de secretina y colecistoquinina del duodeno:
El contenido ácido enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el estómago induce la liberación de secretina.
LosLOSNeisseria aminoácidos y las grasas inducen la liberación de colecistoquinina.
La secretina y la colecistoquinina ingresan alALAmyloidosis torrente sanguíneo y se transportan alALAmyloidosis páncreas.
Estimulación neural:
Estimulación directa por el nervio vago (acetilcolina)
Estimulación a través de otros neurotransmisores:
Péptido intestinal vasoactivo
Péptido liberador de gastrina
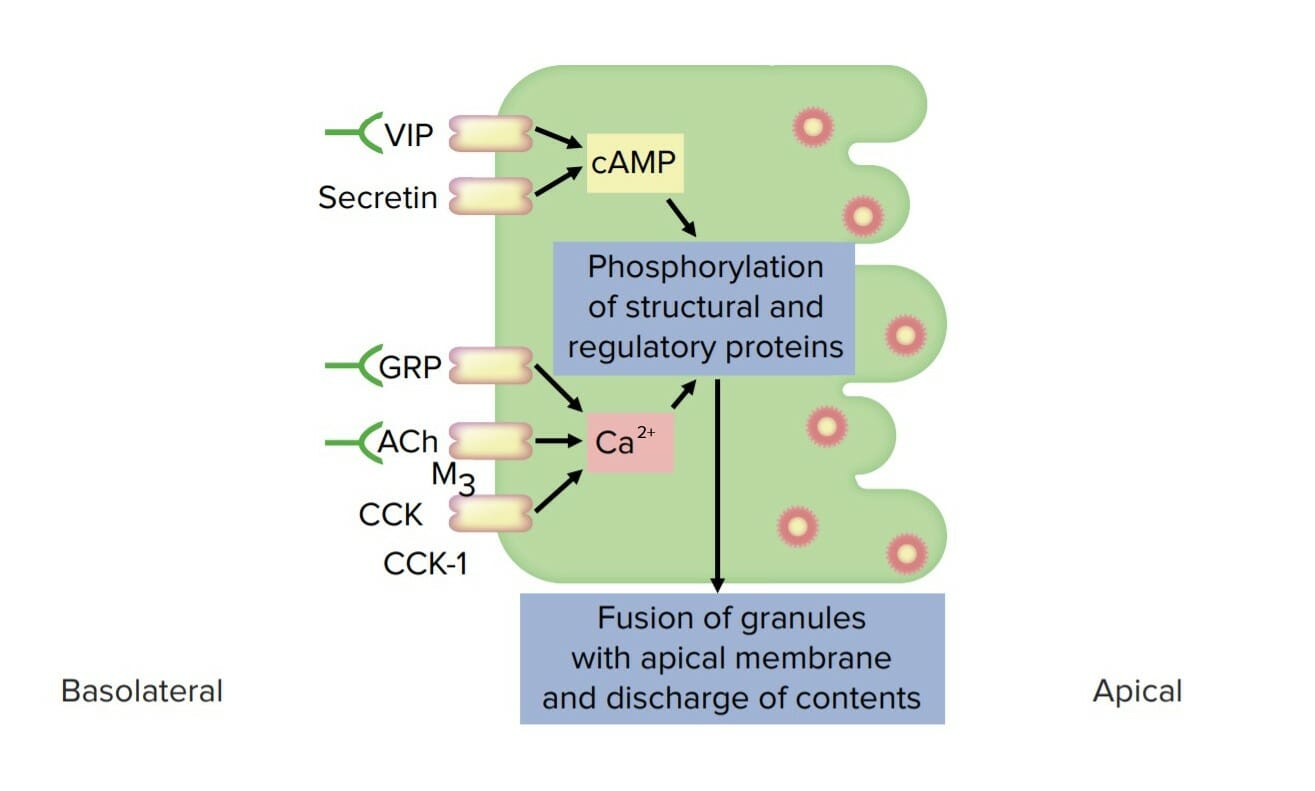
Dos vías intracelulares principales desencadenan la secreción de jugos pancreáticos:
↑ AMPc intracelular, que es causado por:
Secretina
Péptido intestinal vasoactivo
↑ CaCACondylomata acuminata are a clinical manifestation of genital HPV infection. Condylomata acuminata are described as raised, pearly, flesh-colored, papular, cauliflower-like lesions seen in the anogenital region that may cause itching, pain, or bleeding.Condylomata Acuminata (Genital Warts)2+ intracelular:
Acetilcolina
Colecistoquinina
Péptido liberador de gastrina
Ambas vías dan como resultado la fosforilación de proteínas estructurales y reguladoras → inducen el acoplamiento y la fusión de gránulos secretores → las proteínas se secretan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria acinos
Diagrama de una célula pancreática exocrina y sus vías de estimulación de la secreción: Obsérvese cómo el péptido intestinal vasoactivo (VIP) y la secretina aumentan la concentración de cAMP intracelular, mientras que el péptido liberador de gastrina (GRP), la acetilcolina (ACh) y la colecistocinina (CCK) aumentan las concentraciones de Ca2+ intracelular. Ambas vías dan como resultado la fosforilación de las proteínas estructurales y reguladoras, lo que finalmente induce el acoplamiento y la fusión de los gránulos secretores que contienen enzimas.
Imagen por Lecturio.
Relevancia clínica de las secreciones pancreáticas
PancreatitisPancreatitisInflammation of the pancreas. Pancreatitis is classified as acute unless there are computed tomographic or endoscopic retrograde cholangiopancreatographic findings of chronic pancreatitis. The two most common forms of acute pancreatitis are alcoholic pancreatitis and gallstone pancreatitis.Acute Pancreatitis aguda: ocurre cuando las enzimas pancreáticas se retienen dentro del páncreas, lo que resulta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosumla autodigestión. La pancreatitisPancreatitisInflammation of the pancreas. Pancreatitis is classified as acute unless there are computed tomographic or endoscopic retrograde cholangiopancreatographic findings of chronic pancreatitis. The two most common forms of acute pancreatitis are alcoholic pancreatitis and gallstone pancreatitis.Acute Pancreatitis aguda puede ocurrir debido a una obstrucción (e.g., cálculos biliares, cáncer enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la cabeza del páncreas) y puede verse enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum casos de fibrosisFibrosisAny pathological condition where fibrous connective tissue invades any organ, usually as a consequence of inflammation or other injury.Bronchiolitis Obliterans quística.
FibrosisFibrosisAny pathological condition where fibrous connective tissue invades any organ, usually as a consequence of inflammation or other injury.Bronchiolitis Obliterans quística: trastorno autosómico recesivo causado por mutaciones enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el gen CFTR. Las mutaciones conducen a la disfunción de losLOSNeisseria canales de Cl–, lo que resulta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la formación de moco hiperviscoso. El moco hiperviscoso obstruye el flujo de jugo pancreático y bilis enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el tracto gastrointestinal, lo que resulta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum mala digestión, malabsorción y enfermedad hepática y pancreática progresiva. Las presentaciones comunes incluyen infecciones respiratorias crónicas, retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el crecimiento e insuficiencia pancreática (debido a la pérdida de la función exocrina).
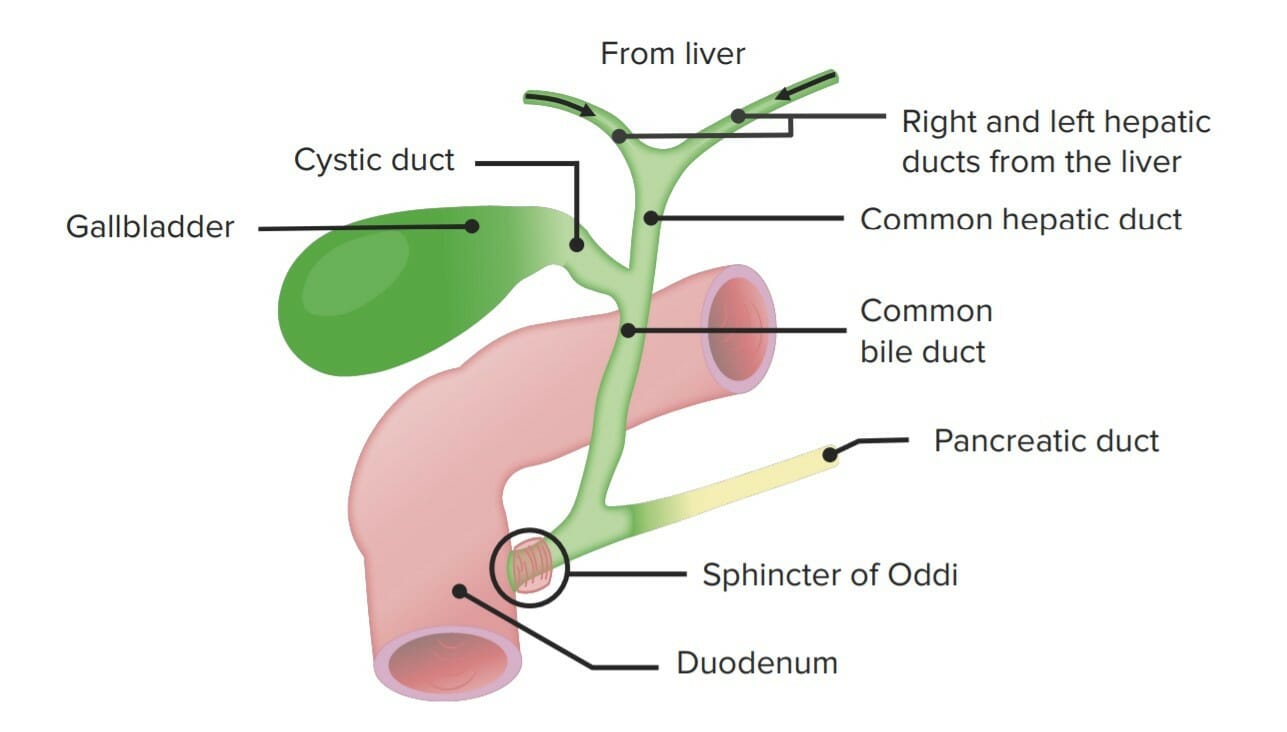
Transportada a la vesícula biliar (a través de losLOSNeisseria conductos hepático y cístico) para su almacenamiento
Función principal: emulsificación (más que digestión) de grasas
Estructura del árbol hepatobiliar
Imagen por Lecturio.
Componentes de las secreciones biliares (i.e., bilis)
Sales biliares
Colesterol
Lecitina
Bilirrubina
Iones
Concentración de bilis enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la vesícula biliar
La función principal de la vesícula biliar es el almacenamiento y la concentración de la bilis. La bilis se puede concentrar enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la vesícula biliar entre 5%‒20% por deshidratación:
Na+:
Reabsorbido desde el lumen a cambio de H+ a través del intercambiador Na+/H+enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana apical
Luego, el Na+ dentro de la célula se bombea a través de la membrana basolateral hacia el espacio intersticial a cambio de K+ a través de la Na+/K+ ATPasa.
Cl–:
Reabsorbido del lumen a cambio de HCO3– a través del intercambiador HCO3–/Cl–enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana apical
El Cl– dentro de la célula se mueve hacia el espacio intersticial a través de losLOSNeisseria canales de Cl– enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la membrana basolateral.
Se mueve desde el lumen de la vesícula biliar hacia el espacio intersticial
A través del movimiento transcelular y paracelular
Mecanismo de concentración biliar: el Na+ se intercambia por H+ en la membrana apical y luego se intercambia por K+ en la membrana basolateral. A continuación, el Cl– se intercambia por HCO3– en la membrana apical y se desplaza al espacio intersticial a través de sus propios canales. El gradiente de concentración que se produce induce el movimiento transcelular y paracelular del agua.
Imagen por Lecturio.
Control y regulación
El hígado produce continuamente bilis; por lo tanto, la regulación se realiza a través de la liberación por la vesícula biliar.
El principal mediador es la colecistoquinina, que tiene 2 efectos principales:
Contracción de la vesícula biliar
Relajación del esfínter de Oddi
Otros reguladores:
Nervio vago: estimulador débil de la contracción de la vesícula biliar
LosLOSNeisseria intestinos secretan alrededor de 1‒2 L de jugo intestinal por día.
Secreciones liberadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el lumen:
Moco
Fluido seroso
Funciones del jugo intestinal:
Protección de las paredes intestinales
Regulación de la función gastrointestinal
Secreciones mínimas para funciones digestivas (i.e., las secreciones contienen muy pocas enzimas)
Secreciones liberadas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la sangre (i.e., moléculas de señalización hormonal):
Freiburghaus, A. U., Roduner, J. B., & Hadorn, H. B. (2021). Activation of human pancreatic proteolytic enzymes: The role of enteropeptidase and trypsin. JPGN Reports, 2(4), e138. https://doi.org/10.1097/PG9.0000000000000138
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Medical Premium le brinda acceso completo a todo el contenido y las funciones
Obtenga Premium para ver todos los vídeos
Verifica tu correo electrónico para obtener una prueba gratuita.
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Premium le ofrece acceso completo a todos los contenidos y funciones, incluido el banco de preguntas de Lecturio con preguntas actualizadas de tipo tablero.