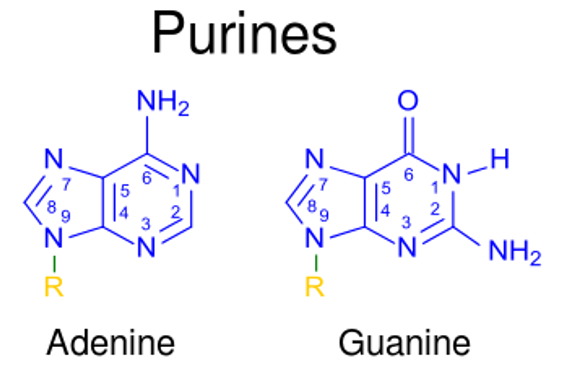
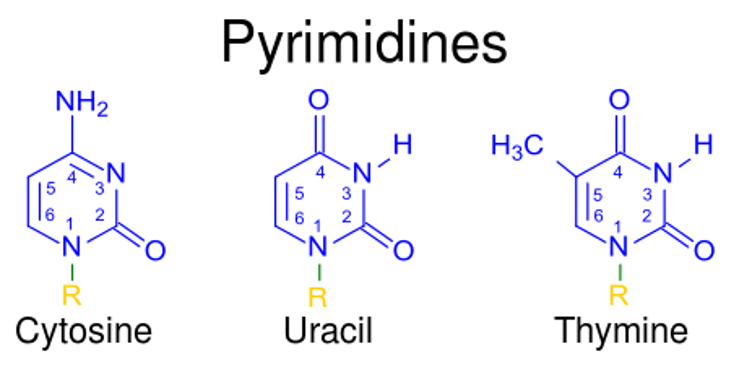
Las purinas y las pirimidinas son compuestos aromáticos heterocíclicos que, junto con losLOSNeisseria grupos azúcar y fosfato, forman losLOSNeisseria componentes importantes de losLOSNeisseria nucleótidos. Las purinas incluyen adenina y guanina, mientras que las pirimidinas incluyen timina (enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ADN), uracilo (enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ARN) y citosina. La síntesis de nucleótidos de purina sigue una serie de reacciones que utilizan donantes de carbono, aminoácidos (e.g., glutamina, aspartato) y bicarbonato. La vía de novo genera inosina monofosfato (IMP), que es el precursor de adenosina monofosfato (AMP) y guanosina monofosfato (GMP). La síntesis de purinas se regula enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria 1ros 2 pasos. La síntesis de nucleótidos de pirimidina también sigue diferentes reacciones, produciendo uridina monofosfato (UMP), que se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum uridina trifosfato (UTP) y citidina trifosfato (CTPCTPPurine and Pyrimidine Metabolism, por sus siglas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum inglés). Para la timina, una parte de losLOSNeisseria desoxirribonucleótidos, se requiere ribonucleósido reductasa para reducir el resto de ribosa. La degradación de losLOSNeisseria nucleótidos da como resultado la producción de xantina y luego de ácido úrico enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum purinas, mientras que las pirimidinas producen losLOSNeisseria aminoácidos, β-alanina y β-aminobutirato.
BasesBasesUsually a hydroxide of lithium, sodium, potassium, rubidium or cesium, but also the carbonates of these metals, ammonia, and the amines.Acid-Base Balance nitrogenadas:
Purinas:
Adenina (A)
Guanina (G)
Pirimidinas:
Timina (T)
Uracilo (U)
Citosina (C)
Otras basesBasesUsually a hydroxide of lithium, sodium, potassium, rubidium or cesium, but also the carbonates of these metals, ammonia, and the amines.Acid-Base Balance menores:
Hipoxantina
Xantina
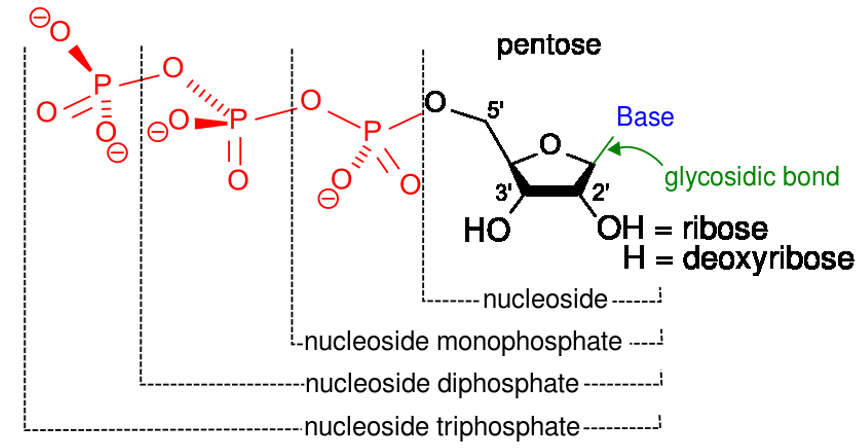
Nucleósidos: 2 componentes:
Una base nitrogenada:
Adenina, guanina, timina y citosina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ADN
Adenina, guanina, uracilo y citosina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ARN
Azúcar pentosa:
Ribosa
Desoxirribosa
Un enlace beta-N-glucosídico une el 1er carbono del azúcar pentosa y N9 de una purina o N1 de una pirimidina (e.g., adenosina, guanosina, citidina, timidina, uridina, inosina).
Nucleótidos: 3 componentes principales:
Base nitrogenada
Azúcar de pentosa
Grupos de fosfato (número variableVariableVariables represent information about something that can change. The design of the measurement scales, or of the methods for obtaining information, will determine the data gathered and the characteristics of that data. As a result, a variable can be qualitative or quantitative, and may be further classified into subgroups.Types of Variables)
Estas moléculas forman el esqueleto del ADN (e.g., adenosina monofosfato, guanosina monofosfato, citidina monofosfato)
> 1 grupo fosfato:
La esterificación de losLOSNeisseria grupos fosfato forma losLOSNeisseria correspondientes nucleósidos difosfatos y trifosfatos (e.g., adenosina trifosfato (ATP), adenosina difosfato (ADP)).
Ácido nucleico:
Polímero de nucleótidos (e.g., ácido ribonucleico (ARN)).
NucleóSido: base + Sugar (azúcar, enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum inglés)
NucleóTido: base + azúcar + fosfaTo
Importancia biomédica
Las principales funciones de losLOSNeisseria nucleótidos:
Componentes básicos de losLOSNeisseria ácidos nucleicos
Actúan como cosustratos y coenzimas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum reacciones bioquímicas
Participan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum las vías de señalización celular y también actúan como segundos mensajeros intracelulares
Proporcionan energía química enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum forma de nucleósidos trifosfatos como el ATP (energía enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum reacciones como la síntesis de aminoácidos, proteínas y membranas celulares)
Síntesis de Purinas
Construcción de la estructura (síntesis de novo)
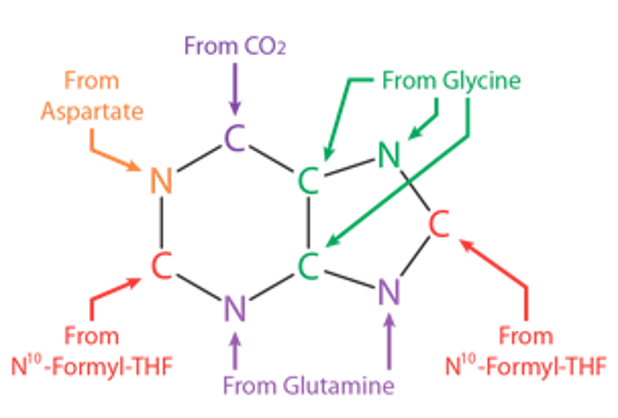
LosLOSNeisseria nucleótidos se forman a partir de moléculas simples: aminoácidos (e.g., glutamina), donantes de carbono (e.g., tetrahidrofolato de formilo) y bicarbonato.
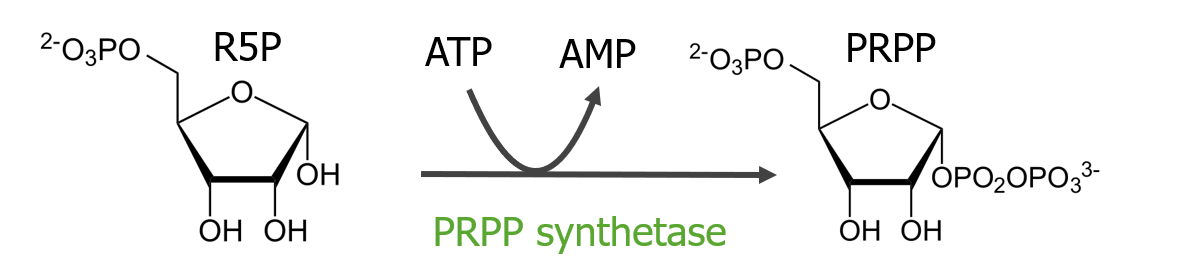
La síntesis de nucleótidos de purina es un proceso de reacción múltiple que comienza con la conversión de ribosa-5-fosfato enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum 5-fosforribosil-1-pirofosfato.
El sitio principal de síntesis es el hígado (intracitoplasmático).
Fuentes atómicas para la síntesis de purinas THF: tetrahidrofolato
Imagen por Lecturio.
Paso 1
Síntesis de 5-fosforribosil-1-pirofosfato
El 5-fosforribosil-1-pirofosfato es el sustrato para la síntesis de purinas.
La ribosa-5-fosfato se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum 5-fosforribosil-1-pirofosfato, con fosfatos provenientes del ATP (reacción que produce AMP).
Correlación clínica: hiperactividad del 5-fosforribosil-1-pirofosfato: trastorno ligado alALAmyloidosis cromosoma X asociado con la sobreproducción de nucleótidos, que se manifiesta con ↑ ácido úrico y anomalías del neurodesarrollo
Síntesis de fosforribosil pirofosfato (PRPP): La ribosa-5-fosfato (R5P) se convierte en PRPP. Los fosfatos provienen del ATP y se produce AMP. La enzima para la conversión es la PRPP sintetasa.
El grupo pirofosfato de 5-fosforribosil-1-pirofosfato se libera enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum esta reacción.
Reacción limitante
Enzima: amidofosforribosiltransferasa
La enzima es inhibida por:
AMP
Guanosina monofosfato (GMP)
Inosina monofosfato (IMP)
Paso 3
Conversión de 5-fosforribosilamina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum ribonucleótido de glicinamida
LosLOSNeisseria pasos posteriores son adiciones para formar un anillo de 5 o 6 componentes.
Se agrega glicina a la 5-fosforribosilamina para formar ribonucleótido de glicinamida.
La glicina aporta el C4, C5 y N7.
Enzima: ribonucleótido de glicinamida sintetasa/fosforribosilamina glicina ligasa
Paso 4
Formilación del ribonucleótido de glicinamidaa ribonucleótido de formilglicinamida
El formiltetrahidrofolato formila el grupo amino del ribonucleótido de glicinamida para formar ribonucleótido de formilglicinamida, aportando el C8 de la purina.
Enzima: ribonucleótido de glicinamida transformilasa/fosforribosil glicinamida formiltransferasa
Paso 5
Conversión del ribonucleótido de formilglicinamidaa ribonucleótido de formilglicinamidina
EnENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum esta reacción dependiente de adenosina trifosfato (ATP), la glutamina dona el N3, formando ribonucleótido de formilglicinamidina.
Enzima: ribonucleótido de formilglicinamidina sintetasa/fosforribosil formil glicinamida sintasa
Paso 6
Formación del anillo de purina imidazol
Esta es una reacción dependiente de ATP que conduce a la formación y cierre del anillo de purina.
El ribonucleótido de 5-aminoimidazol se forma a partir de esta reacción.
Enzima: ribonucleótido de 5-aminoimidazol sintetasa/fosforribosil formil glicinamida cicloligasa
Paso 7
Carboxilación del ribonucleótido de 5-aminoimidazol
Esta es una carboxilación dependiente de ATP del ribonucleótido de 5-aminoimidazol a ribonucleótido de carboxiaminoimidazol, enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum presencia de bicarbonato
El C6 de la purina lo aporta el bicarbonato.
Enzima: ribonucleótido de 5-aminoimidazol carboxilasa
Paso 8
Formación del ribonucleótido de 5-aminoimidazol-4-(N-succinilcarboxamida)
La adición de aspartato forma un enlace amida con C6 para formar ribonucleótido de 5-aminoimidazol-4.
El N1 de la purina es aportado por el aspartato.
Enzima: ribonucleótido de 5-aminoimidazol-4- sintetasa
Paso 9
Eliminación de fumarato
El ribonucleótido de 5-aminoimidazol-4-carboxamida se forma por la escisión del grupo fumarato.
Formilación para formar el ribonucleótido de 5-formaminoimidazol-4-carboxamida
La formilación ocurre por reacción entre el grupo amino del 5-aminoimidazol-4-carboxamida y N10-formil tetrahidrofolato para formar ribonucleótido de 5-formaminoimidazol-4-carboxamida.
El C2 del anillo de purina lo aporta el tetrahidrofolato de N10-formilo.
La inosina monofosfato se forma por el cierre enzimático del anillo mayor del ribonucleótido de 5-formaminoimidazol-4-carboxamida con liberación de agua.
La inosina monofosfato es el precursor de AMP y GMP.
AICAR: ribonucleótido de 5-aminoimidazol-4-carboxamida (por sus siglas en inglés)
AIR: ribonucleótido de 5-aminoimidazol (por sus siglas en inglés)
FGAM: ribonucleótido de formilglicinamidina (por sus siglas en inglés)
FGAR: ribonucleótido de formilglicinamida (por sus siglas en inglés)
GAR: ribonucleótido de glicinamida (por sus siglas en inglés)
IMP: inosina monofosfato
PRPP: pirofosfato de fosforribosil (por sus siglas en inglés)
PRA: 5-fosforribosilamina
SAICAR: ribonucleótido de 5-aminoimidazol-4-(N-succinilcarboxamida) (por sus siglas en inglés)
THF: tetrahidrofolato
Papel del folato
El ácido fólico se compone de ácido p-aminobenzoico, glutamina y pteridina y está disponible para su uso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum su forma activa: ácido tetrahidrofólico.
La falta de folato conduce a una disminución de la síntesis de nucleótidos.
2 consecuencias importantes de la deficiencia de ácido fólico son la anemiaAnemiaAnemia is a condition in which individuals have low Hb levels, which can arise from various causes. Anemia is accompanied by a reduced number of RBCs and may manifest with fatigue, shortness of breath, pallor, and weakness. Subtypes are classified by the size of RBCs, chronicity, and etiology. Anemia: Overview and Types megaloblástica y la espina bífida enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria recién nacidos (debido a la deficiencia materna de folato).
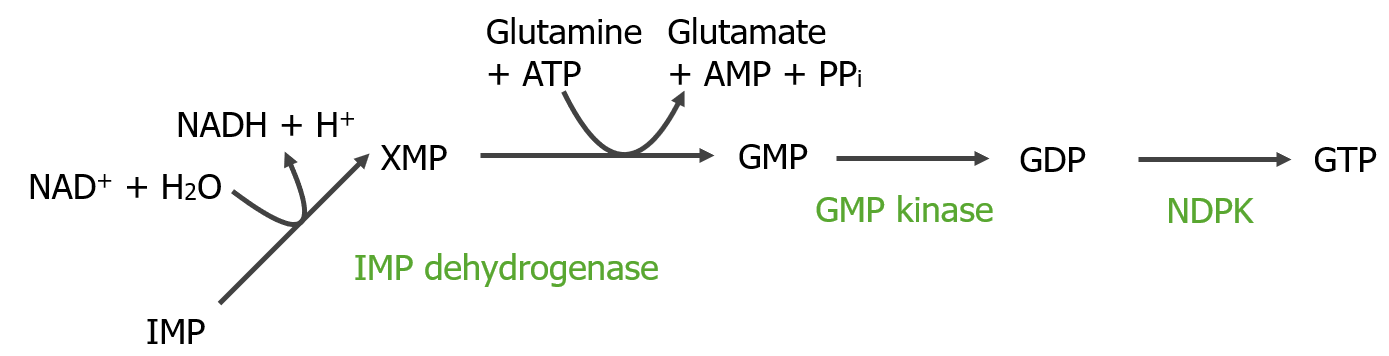
La inosina monofosfato se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum adenina y guanina como AMP y GMP. Formado a partir de GMP, la guanosina trifosfato (GTP) proporciona la energía para convertir IMP enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum AMP.
Se liberan iones H+ (y son aceptados por el NADNAD+A coenzyme composed of ribosylnicotinamide 5′-diphosphate coupled to adenosine 5′-phosphate by pyrophosphate linkage. It is found widely in nature and is involved in numerous enzymatic reactions in which it serves as an electron carrier by being alternately oxidized (NAD+) and reduced (NADH). Pentose Phosphate Pathway+).
El micofenolato, un inmunosupresor, inhibe la IMP deshidrogenasa, reduciendo la proliferación de células inmunitarias.
Conversión de IMP a GMP y luego a GTP: NAD+: nicotinamida adenina dinucleótido (oxidado) NADH: nicotinamida adenina dinucleótido (reducido) NDPK: nucleósido difosfato quinasa (por sus siglas en inglés) PPi: pirofosfato
Imagen por Lecturio.
Síntesis de AMP
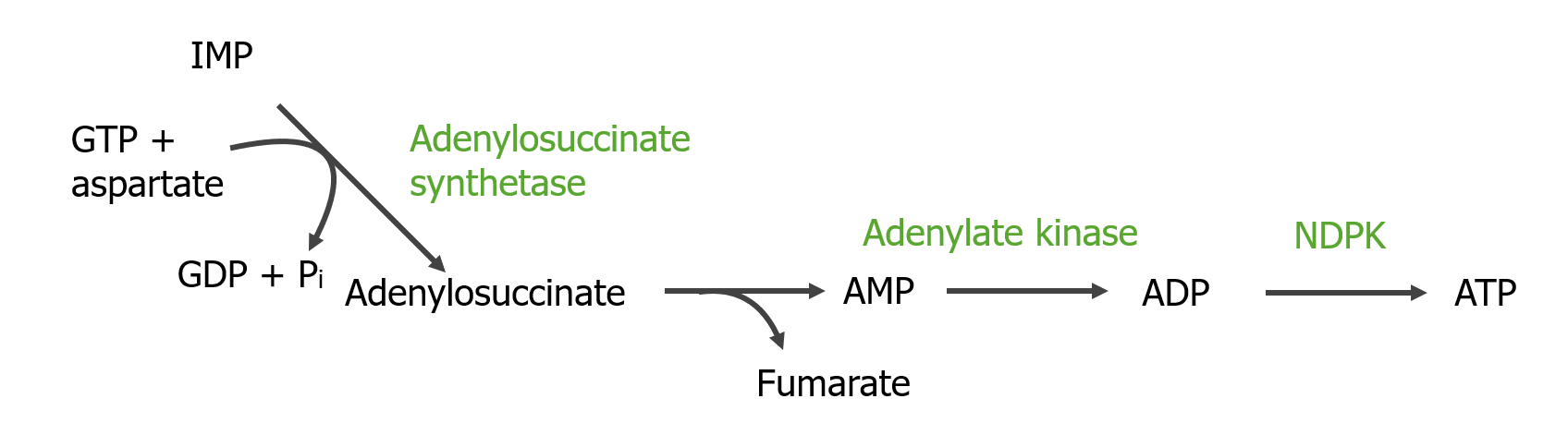
Paso 1: donación del grupo amino por aspartato
El grupo amino del aspartato (enlaces a IMP) + hidrólisis de GTP → adenilosuccinato
Enzima: adenilosuccinato sintetasa
Paso 2: eliminación de fumarato para formar AMP
El adenilosuccinato se convierte enzimáticamente enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum AMP mediante la eliminación del fumarato.
Enzima: adenilosuccinasa/adenilosuccinato liasa
Conversión de IMP a AMP y luego a ATP: NDPK: nucleósido difosfato quinasa (por sus siglas en inglés) Pi: fosfato inorgánico
Imagen por Lecturio.
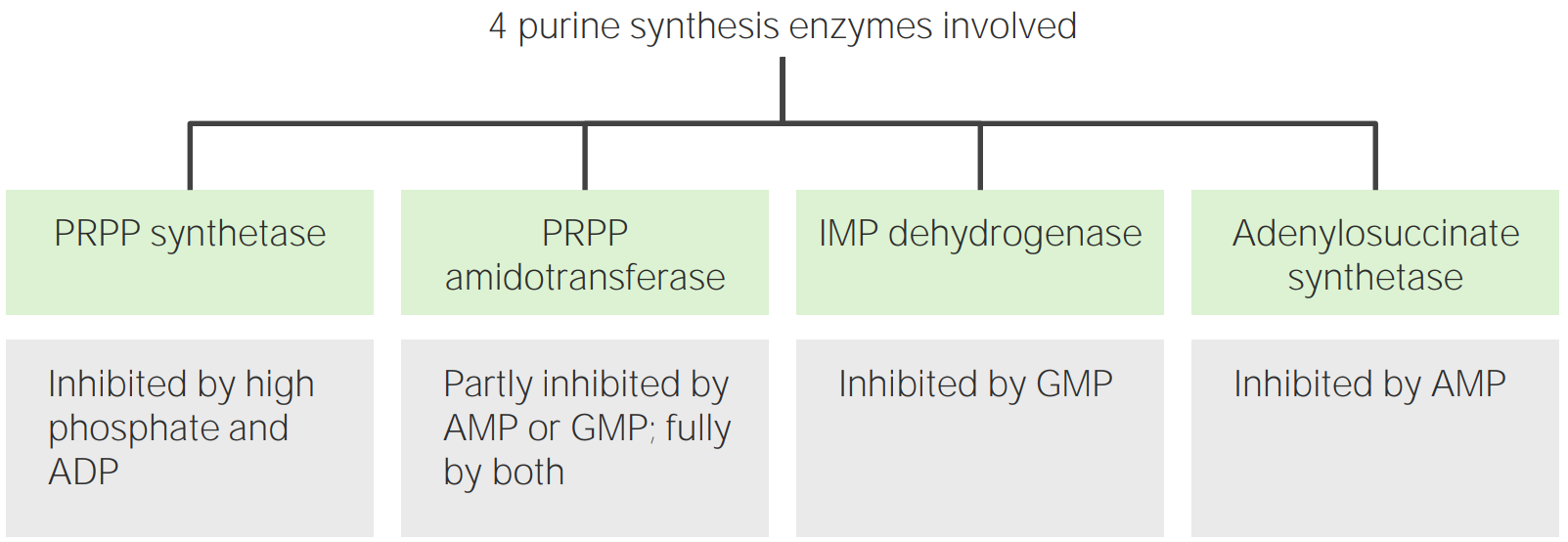
Regulación de la síntesis
La síntesis de IMP, ATP y GTP está regulada para controlar la cantidad de nucleótidos de purina producidos.
La enzima 5-fosforribosil-1-pirofosfato sintetasa (paso 1) es inhibida por ADP y GDP.
La enzima amidofosforribosiltransferasa (paso 2) es inhibida por:
AMP
GMP
IMP
La enzima adenilosuccinato sintetasa (síntesis de AMP) es inhibida por el AMP.
La enzima IMP deshidrogenasa (enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la síntesis de GMP) es inhibida por el GMP.
LosLOSNeisseria factores externos que afectan la síntesis de purinas incluyen análogos de purinas:
Tiopurinas (inhiben la síntesis de novo de purinas)
6-Mercaptopurina: agente antineoplásico e inmunosupresor
6-Tioguanina
Azatioprina (inmunosupresor): se somete a una reducción no enzimática a 6-mercaptopurina
Generación de nucleótidos a partir de la descomposición de ácidos nucleicos
Las purinas libres se vuelven a convertir enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum sus respectivos nucleótidos a través de vías de reciclaje.
El 5-fosforribosil-1-pirofosfato es un componente esencial enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum esta vía.
Correlación clínica: síndrome de Lesch-Nyhan: trastorno recesivo ligado alALAmyloidosis cromosoma X causado por un defecto enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la hipoxantina-guanina fosforribosiltransferasa (incapaz de reciclar las basesBasesUsually a hydroxide of lithium, sodium, potassium, rubidium or cesium, but also the carbonates of these metals, ammonia, and the amines.Acid-Base Balance de purina → ↑ ácido úrico)
La vía de reciclaje que reutiliza nucleótidos para su utilización
Imagen por Lecturio.
Importancia
EnENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum tejidos como losLOSNeisseria eritrocitos y el cerebro, la vía de reciclaje es importante debido a la ausencia de síntesis de purinas de novo.
La vía economiza el gasto de energía intracelular.
Catabolismo de Nucleótidos de Purina
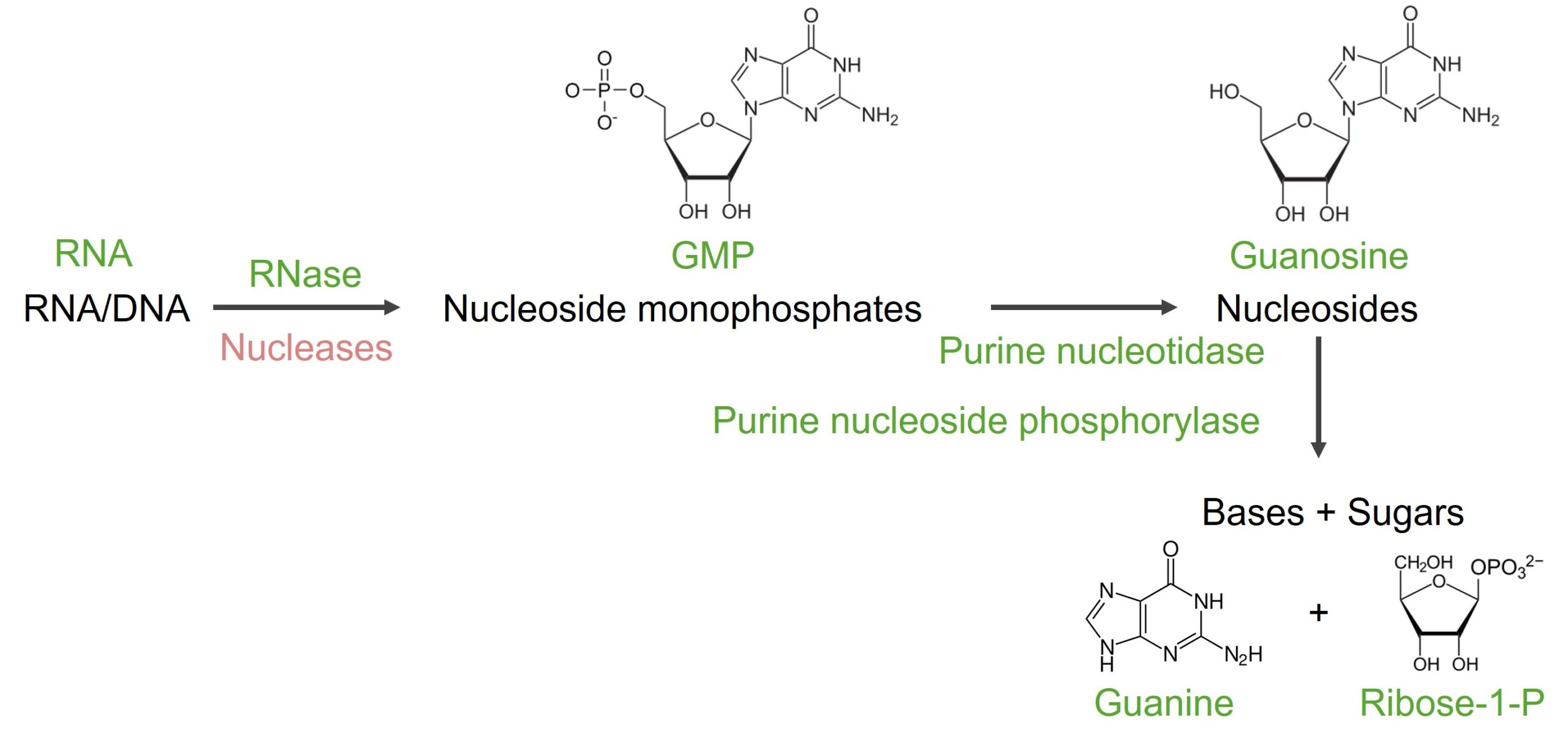
El ácido nucleico (ARN/ADN) se descompone enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum nucleótidos mediante nucleasas. Para degradar losLOSNeisseria nucleótidos de purina, primero se eliminan el fosfato y la ribosa, y otras reacciones conducen a la xantina y luego alALAmyloidosis ácido úrico.
Guanosina monofosfato
Conversión de nucleótido a nucleósido (GMP a guanosina) por la enzima nucleotidasa, lo que resulta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la eliminación de fosfato
La guanosina se descompone aún más:
La reacción conduce a guanina y ribosa-1-fosfato.
Enzima: purina nucleósido fosforilasa
La desaminación de la guanina conduce a la formación de xantina.
Degradación de guanina
Imagen por Lecturio.
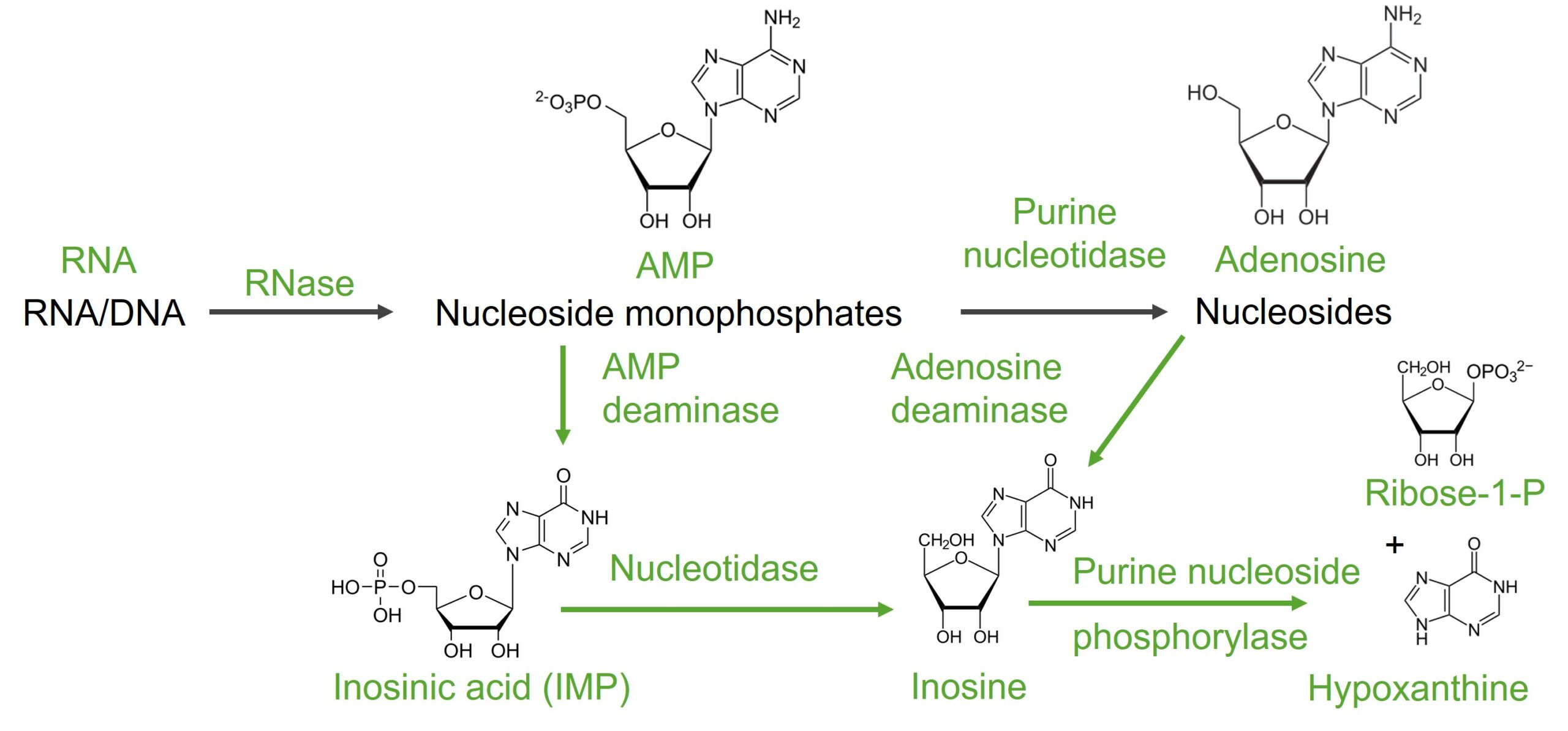
AMP
La conversión de ácidos nucleicos (ARN/ADN a AMP a basesBasesUsually a hydroxide of lithium, sodium, potassium, rubidium or cesium, but also the carbonates of these metals, ammonia, and the amines.Acid-Base Balance) puede tener diferentes vías, utilizando diferentes desaminasas.
1era vía:
AMP → adenosina: catalizada por la enzima purina nucleotidasa, con eliminación del fosfato
La adenosina se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum inosina por la adenosina desaminasa
La inosina es degradada por la purina nucleósido fosforilasa a hipoxantina y ribosa-1-fosfato.
La hipoxantina es oxidada a xantina por la xantina oxidasa.
2da vía:
AMP → ácido inosínico o IMP: catalizada por la AMP desaminasa
El IMP se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum inosina por la nucleotidasa.
La inosina es degradada por la purina nucleósido fosforilasa a hipoxantina y ribosa-1-fosfato.
La hipoxantina es oxidada a xantina por la xantina oxidasa.
Correlación clínica:
Deficiencia de adenosina desaminasa: conduce a ↑ desoxi-ATP, desoxi-GTP (tóxico para las células inmunitarias como losLOSNeisseria linfocitos T)
Deficiencia de purina nucleósido fosforilasa:conduce a ↑ desoxi-ATP, desoxi-GTP (tóxico para las células inmunitarias como losLOSNeisseria linfocitos T) y también se asocia con retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el desarrollo
Degradación de la adenina
Imagen por Lecturio.
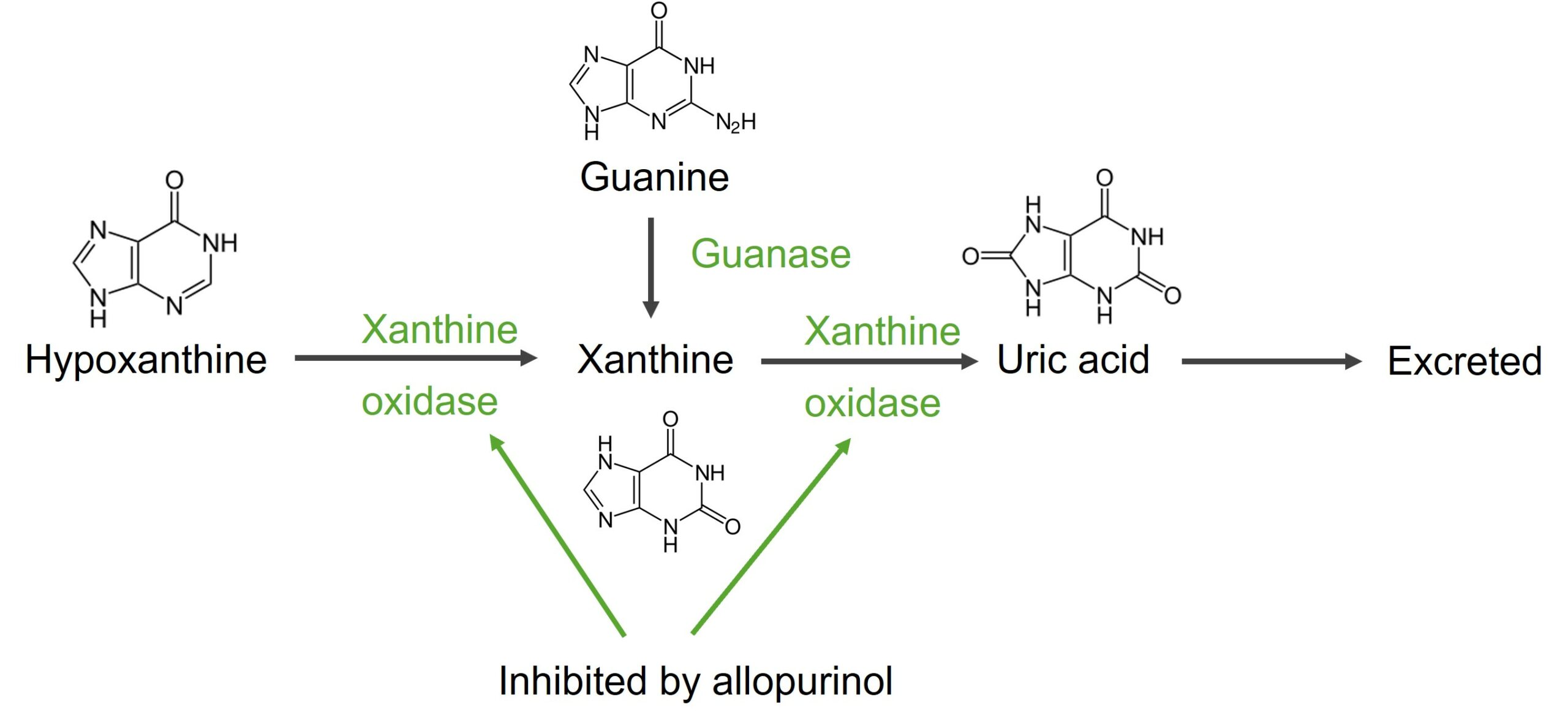
Xantina
Tanto la adenosina como la guanosina se convierten enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum xantina.
Adenosina → inosina → hipoxantina → xantina
Guanosina → guanina → xantina
Xantina oxidasa:
Cataliza la reacción de hipoxantina a xantina y de xantina a ácido úrico
El producto final, el ácido úrico, se excreta enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la orina.
Correlación clínica: el alopurinol, un inhibidor de la xantina oxidasa, se utiliza para el tratamiento de la gota.
Degradación de guanina e hipoxantina en ácido úrico
La base de pirimidina se sintetiza primero y luego se incorpora alALAmyloidosis nucleótido (el anillo se completa antes de unirse a la ribosa-5-fosfato).
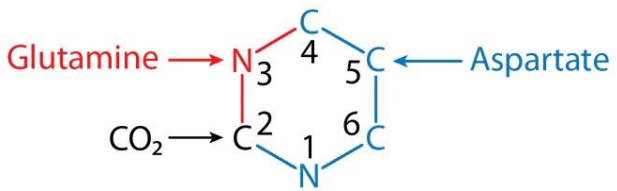
Fuentes de losLOSNeisseria átomos de carbono y nitrógeno de la pirimidina:
La glutamina y el bicarbonato aportan N3 y C2, respectivamente, que se combinan para formar carbamoil fosfato.
El aspartato aporta N1, C6, C5 y C4
Fuentes de los átomos de carbono y nitrógeno en la síntesis de pirimidinas
Imagen por Lecturio.
Paso 1
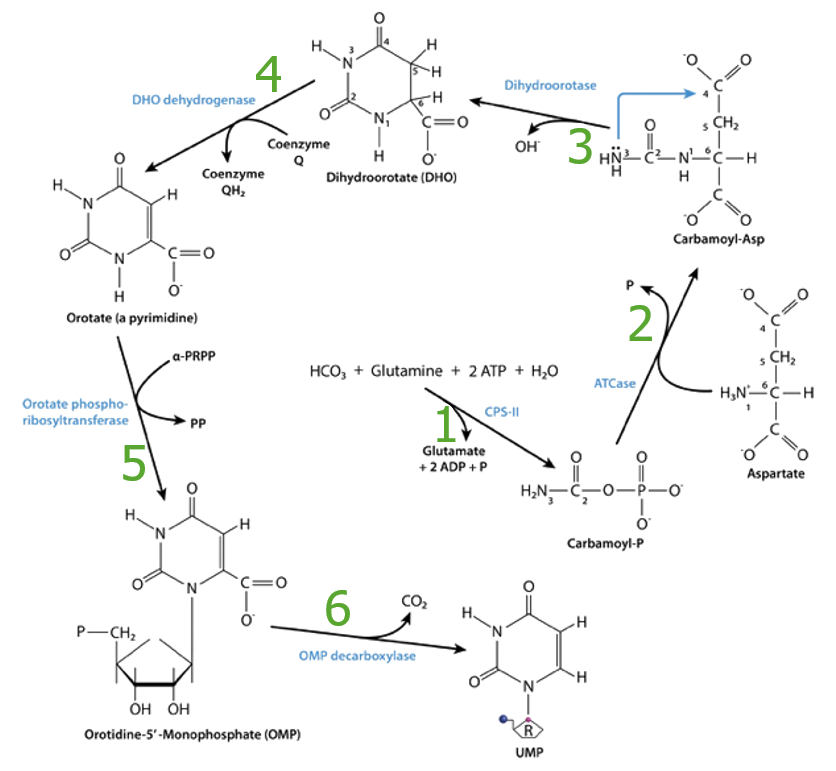
Síntesis de carbamoil fosfato
Esta reacción ocurre enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el citoplasma.
El nitrógeno de la glutamina y el carbono del bicarbonato reaccionan para formar carbamoil fosfato.
Enzima: carbamoil fosfato sintetasa II
Paso 2
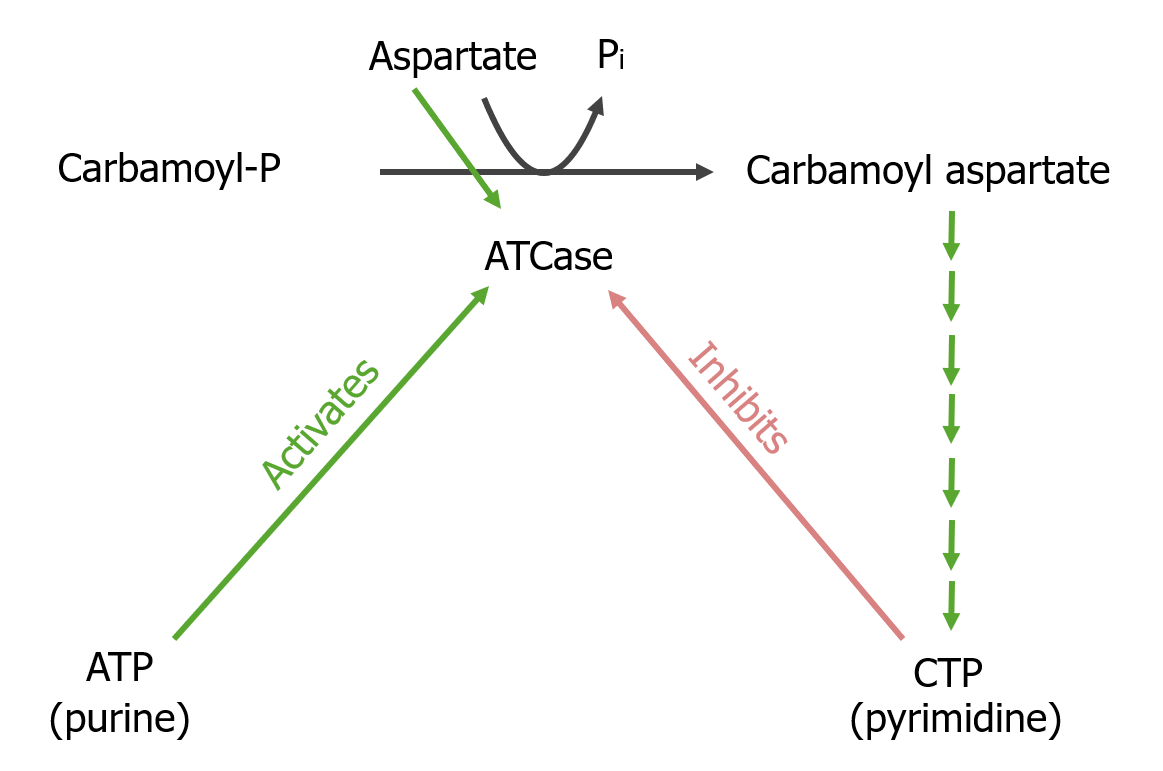
Síntesis de carbamoil aspartato
Reacción limitante
El carbamoil fosfato reacciona con el aspartato para producir carbamoil aspartato.
LosLOSNeisseria átomos C2 y N3 derivan del carbamoil fosfato.
Reacción limitante de la síntesis de pirimidinas: La reacción convierte al carbamoil fosfato en carbamoil aspartato, catalizada por la aspartil transcarbamoilasa (ATCase, por sus siglas en inglés). Las reacciones posteriores finalmente conducen al producto final, la citidina trifosfato (CTP). La aspartil transcarbamoilasa es activada por ATP e inhibida por CTP.
Imagen por Lecturio.
Paso 3
Formación del anillo de pirimidina
Se elimina una molécula de agua y el carbamoil aspartato se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum un compuesto enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum forma de anillo (dihidroorotato).
Enzima: dihidroorotasa
Paso 4
Oxidación del dihidroorotato
La eliminación de átomos de hidrógeno (deshidrogenación) de las posiciones C5 y C6 produce ácido orótico.
Enzima: dihidroorotato deshidrogenasa
Coenzima: NADNAD+A coenzyme composed of ribosylnicotinamide 5′-diphosphate coupled to adenosine 5′-phosphate by pyrophosphate linkage. It is found widely in nature and is involved in numerous enzymatic reactions in which it serves as an electron carrier by being alternately oxidized (NAD+) and reduced (NADH). Pentose Phosphate Pathway
Ácido orótico + ribosa-5-fosfato → orotidina monofosfato o ácido orotidílico
El 5-fosforribosil-1-pirofosfato es el donante de ribosa-5-fosfato.
Enzima: orotato fosforribosiltransferasa
Paso 6
Descarboxilación para formar monofosfato de uridina (UMP)
La orotidina monofosfato sufre una descarboxilación.
La uridina monofosfato (UMP) se produce por la eliminación de C1 enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum forma de CO2, lo que convierte a la uridina enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la primera pirimidina que se sintetiza.
Nota: las últimas 2 enzimas de esta vía, la orotato fosforribosiltransferasa y la OMPOMPPurine and Pyrimidine Metabolism descarboxilasa, se encuentran enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el mismo polipéptido, UMP sintasa. La UMP sintasa cataliza la conversión del ácido orótico enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum UMP.
Tabla: Resumen de la síntesis de novo de pirimidinas
*cataliza la reacción limitante
OMP: orotidina-5-monofosfato
Resumen de la síntesis de pirimidinas, enzimas: 1. CPS II: carbamoil fosfato sintetasa II 2. ATCase: aspartil transcarbamoilasa (por sus siglas en inglés) 3. Dihidroorotasa 4. Dihidroorotato (DHO) deshidrogenasa 5. Orotato fosforribosiltransferasa 6. Orotidina-5-monofosfato (OMP) descarboxilasa
Imagen por Lecturio.
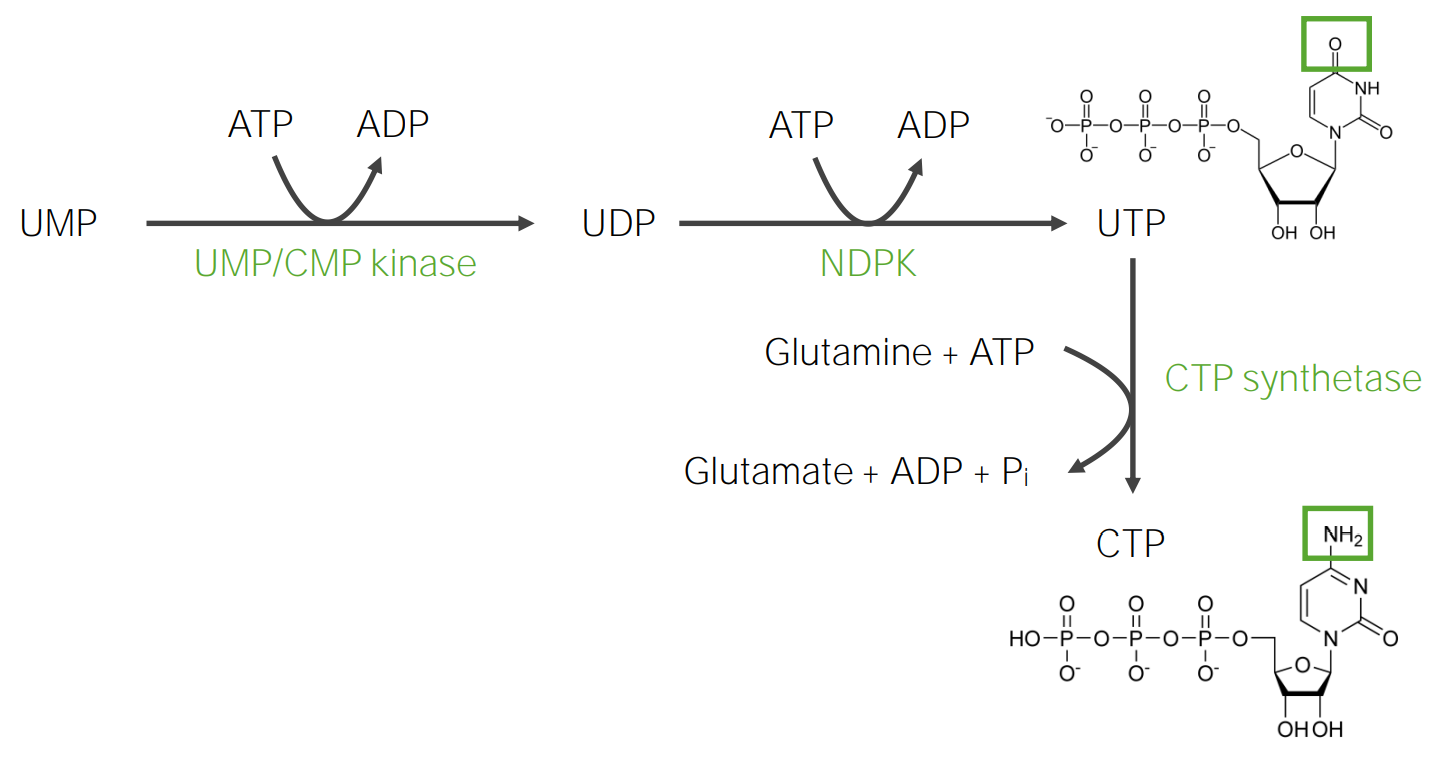
Síntesis de la uridina trifosfato y la citidina trifosfato
El UTP y el CTPCTPPurine and Pyrimidine Metabolism se utilizan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la síntesis de ARN.
UTP:
Paso 1:
La fosforilación de UMP por el ATP produce uridina difosfato (UDP)
El UTP se convierte enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumCTPCTPPurine and Pyrimidine Metabolism (citidina trifosfato) mediante la adición de un grupo amino de la glutamina.
El ADN es diferente del ARN, ya que el ADN tiene desoxirribosa, enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum lugar de ribosa, y timina (5-metiluracilo), enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum lugar de uracilo.
LosLOSNeisseria desoxirribonucleótidos se generan a partir de sus correspondientes ribonucleótidos.
Las ribonucleótido reductasas reducen losLOSNeisseria ribonucleósidos difosfatos a desoxirribonucleósidos difosfatos.
LosLOSNeisseria desoxirribonucleósidos difosfatos, a su vez, se convierten enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum desoxirribonucleósido trifosfato mediante la nucleósido difosfato quinasa.
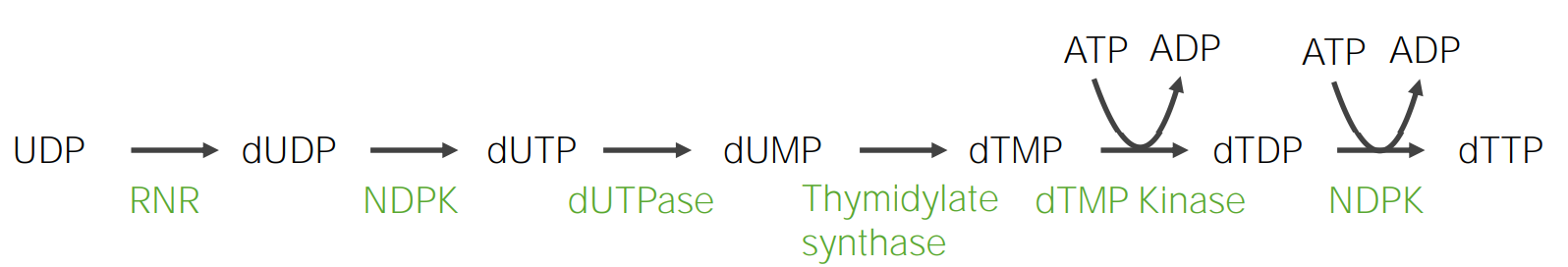
La timina es una pirimidina presente enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ADN; por lo tanto, la ribosa del nucleótido correspondiente requiere reducción.
Paso 1:
UDP → desoxiuridina difosfato (dUDP)
Enzima: ribonucleótido reductasa
Paso 2:
dUDP → desoxiuridina trifosfato (dUTP)
Enzima: nucleósido difosfato quinasa
Paso 3:
dUTP → desoxiuridina monofosfato (dUMP)
Enzima: dUTP difosfohidrolasa
Paso 4:
dUMP se metila a desoxitimidina monofosfato (dTMP).
Enzima: timidilato sintasa
Requiere de metileno tetrahidrofolato (como donador de metilo)
Paso 5:
dTMP se fosforila (por ATP) a desoxitimidina trifosfato (dTTP).
La fosforilación ocurre enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum 2 rondas.
Correlación clínica: 5-fluorouracilo: agente antimetabolito (usado enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el cáncer) que inhibe la timidilato sintasa y disminuye la síntesis de ADN
Formación de timina en forma de desoxitimidina trifosfato (dTTP) dTDP: desoxitimidina difosfato dTMP: desoxitimidina monofosfato dTTP: desoxitimidina trifosfato dUDP: desoxiuridina difosfato dUMP: desoxiuridina monofosfato dUTPasa: desoxiuridina trifosfatasa NDPK: nucleósido difosfato quinasa (por sus siglas en inglés) RNR: ribonucleótido reductasa UDP: difosfato de uridina
Imagen por Lecturio.
Regulación de la síntesis
La enzima carbamoil fosfato sintetasa II enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el paso 1:
Activada por 5-fosforribosil-1-pirofosfato y ATP
Inhibida por UTP y UDP
La enzima aspartil transcarbamoilasa en el paso 2 es inhibida alostéricamente por CTP.
La enzima OMP descarboxilasa (paso 6) es inhibida por UMP.
LosLOSNeisseria factores externos incluyen análogos de pirimidinas (utilizados como agentes antineoplásicos):
5-Fluorouracilo
Capecitabina
Citarabina
Gemcitabina
Vía de reciclaje de nucleótidos de pirimidina
AlALAmyloidosis igual que las purinas, las pirimidinas se reciclan a partir de losLOSNeisseria productos intermedios derivados de losLOSNeisseria ácidos nucleicos.
Las reacciones convierten losLOSNeisseria ribonucleósidos (uridina, citidina) y losLOSNeisseria desoxirribonucleósidos (timidina, desoxicitidina) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum nucleótidos.
Las quinasas o fosforiltransferasascatalizan la transferencia de grupos fosforilo (del ATP) a losLOSNeisseria difosfatos, produciendo trifosfatos:
Ribonucleósidos difosfatos + ATP → ribonucleósido trifosfato + ADP
Desoxirribonucleósidos difosfatos + ATP → desoxirribonucleósido trifosfato + ADP
Las células animales descomponen losLOSNeisseria nucleótidos de pirimidinas enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumbasesBasesUsually a hydroxide of lithium, sodium, potassium, rubidium or cesium, but also the carbonates of these metals, ammonia, and the amines.Acid-Base Balance nitrogenadas, y el uracilo y la timina resultantes se degradan (a través de reducción) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el hígado.
AlALAmyloidosis igual que enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema NodosumlosLOSNeisseria nucleótidos de purinas, las nucleasas degradan el ácido nucleico (ARN/ADN) enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum nucleótidos.
La citosina esdegradada a uracilo por la eliminación de un grupo amino.
Luego, tanto el uracilo como la timina se reducen a dihidrouracilo y dihidrotimina, respectivamente, que reaccionan hasta losLOSNeisseria productos finales:
El β-aminobutirato y la β-alanina se utilizan además enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el metabolismo de losLOSNeisseria aminoácidos.
LosLOSNeisseria iones de amonio (NH4+) liberados por la descomposición se utilizan enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el ciclo de la ureaUreaA compound formed in the liver from ammonia produced by the deamination of amino acids. It is the principal end product of protein catabolism and constitutes about one half of the total urinary solids.Urea Cycle.
Degradación de uracilo y timina NADPH: nicotinamida adenina dinucleótido fosfato
Falta de enzima → defectos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la vía de reciclaje de purinas
Pubertad retrasada
Automutilación
Retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el desarrollo
Deterioro de la función renal
SCIDSCIDSevere combined immunodeficiency (SCID), also called “bubble boy disease,” is a rare genetic disorder in which the development of functional B and T cells is disturbed due to several genetic mutations that result in reduced or absent immune function.Severe Combined Immunodeficiency (SCID)
Infecciones a repetición, abscesos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la capa profunda de la piel o enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum órganos
CandidiasisCandidiasisCandida is a genus of dimorphic, opportunistic fungi. Candida albicans is part of the normal human flora and is the most common cause of candidiasis. The clinical presentation varies and can include localized mucocutaneous infections (e.g., oropharyngeal, esophageal, intertriginous, and vulvovaginal candidiasis) and invasive disease (e.g., candidemia, intraabdominal abscess, pericarditis, and meningitis). Candida/Candidiasis mucocutánea
Retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el crecimiento
Litiasis renal
↓ APRT
Mutación autosómica recesiva → defectos enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum la vía de reciclaje de purinas
Cólico renal
Infecciones urinarias recurrentes
Náuseas
Vómitos
Xantinuria
↓ Xantina oxidasa
Hipouricemia
Nefrolitiasis
Lesión renal aguda
ADA: adenosina desaminasa
APRT: adenina fosforribosiltransferasa
HGPRT: hipoxantina guanina fosforribosiltransferasa
PRPP: pirofosfato de fosforribosil (por sus siglas en inglés)
SCID: inmunodeficiencia combinada severa (por sus siglas en inglés)
Tabla: Trastornos del metabolismo de las pirimidinas
Retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el crecimiento
Retraso enENErythema nodosum is an immune-mediated panniculitis (inflammation of the subcutaneous fat) caused by a type IV (delayed-type) hypersensitivity reaction. It commonly manifests in young women as tender, erythematous nodules on the shins.Erythema Nodosum el desarrollo
AnemiaAnemiaAnemia is a condition in which individuals have low Hb levels, which can arise from various causes. Anemia is accompanied by a reduced number of RBCs and may manifest with fatigue, shortness of breath, pallor, and weakness. Subtypes are classified by the size of RBCs, chronicity, and etiology. Anemia: Overview and Types megaloblástica
Del Arco, J., Ibáñez, F. J., Acosta, J., & Fernández-Lucas, J. (2024). Biotechnological applications of purine and pyrimidine deaminases. Biotechnology Advances, 77, 108473. https://doi.org/10.1016/j.biotechadv.2024.108473
Hernandez, D. M., Lopatkin, A. J., Palmer, A. C., & Manson, A. L. (2024). Purine and pyrimidine synthesis differently affect the strength of the inoculum effect for aminoglycoside and β-lactam antibiotics. Microbiology Spectrum, 12(12), e01895-24. https://doi.org/10.1128/spectrum.01895-24
Jurecka, A., & Tylki-Szymańska, A. (2022). Inborn errors of purine and pyrimidine metabolism: A guide to diagnosis. Molecular Genetics and Metabolism, 136(3), 164–176. https://doi.org/10.1016/j.ymgme.2022.02.007
Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V. W., & Weil, P. A. (2022). Harper’s illustrated biochemistry (33rd ed.). McGraw-Hill Education.
Nassogne, M. C., Marie, S., & Dewulf, J. P. (2024). Neurological presentations of inborn errors of purine and pyrimidine metabolism. European Journal of Paediatric Neurology, 48, 69–77. https://doi.org/10.1016/j.ejpn.2023.11.013
Nelson, D. L., & Cox, M. M. (2021). Lehninger principles of biochemistry (8th ed.). W. H. Freeman.
Ragab, A. (2025). Recent advances in the synthesis, reaction, and bio-evaluation potential of purines as precursor pharmacophores in chemical reactions: A review. RSC Advances, 15(5), 3607–3645. https://doi.org/10.1039/D4RA08271K
Suleiman, H., Naja, M., Fares, D., Abou-Kheir, W., & El-Sitt, S. (2024). Harnessing nucleotide metabolism and immunity in cancer: A tumour microenvironment perspective. The FEBS Journal. https://doi.org/10.1111/febs.17278
¡Crea tu cuenta gratis o inicia una sesión para seguir leyendo!
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Medical Premium le brinda acceso completo a todo el contenido y las funciones
Obtenga Premium para ver todos los vídeos
Verifica tu correo electrónico para obtener una prueba gratuita.
Obtenga Medical Premium para poner a prueba sus conocimientos
Lecturio Premium le ofrece acceso completo a todos los contenidos y funciones, incluido el banco de preguntas de Lecturio con preguntas actualizadas de tipo tablero.