Playlist
Show Playlist
Hide Playlist
T-Cell Development in Thymus and Genetic Recombination of TCR – Lymphocyte Development

-
Slides Adaptive Immune System.pdf
-
Reference List Immune System.pdf
-
Download Lecture Overview
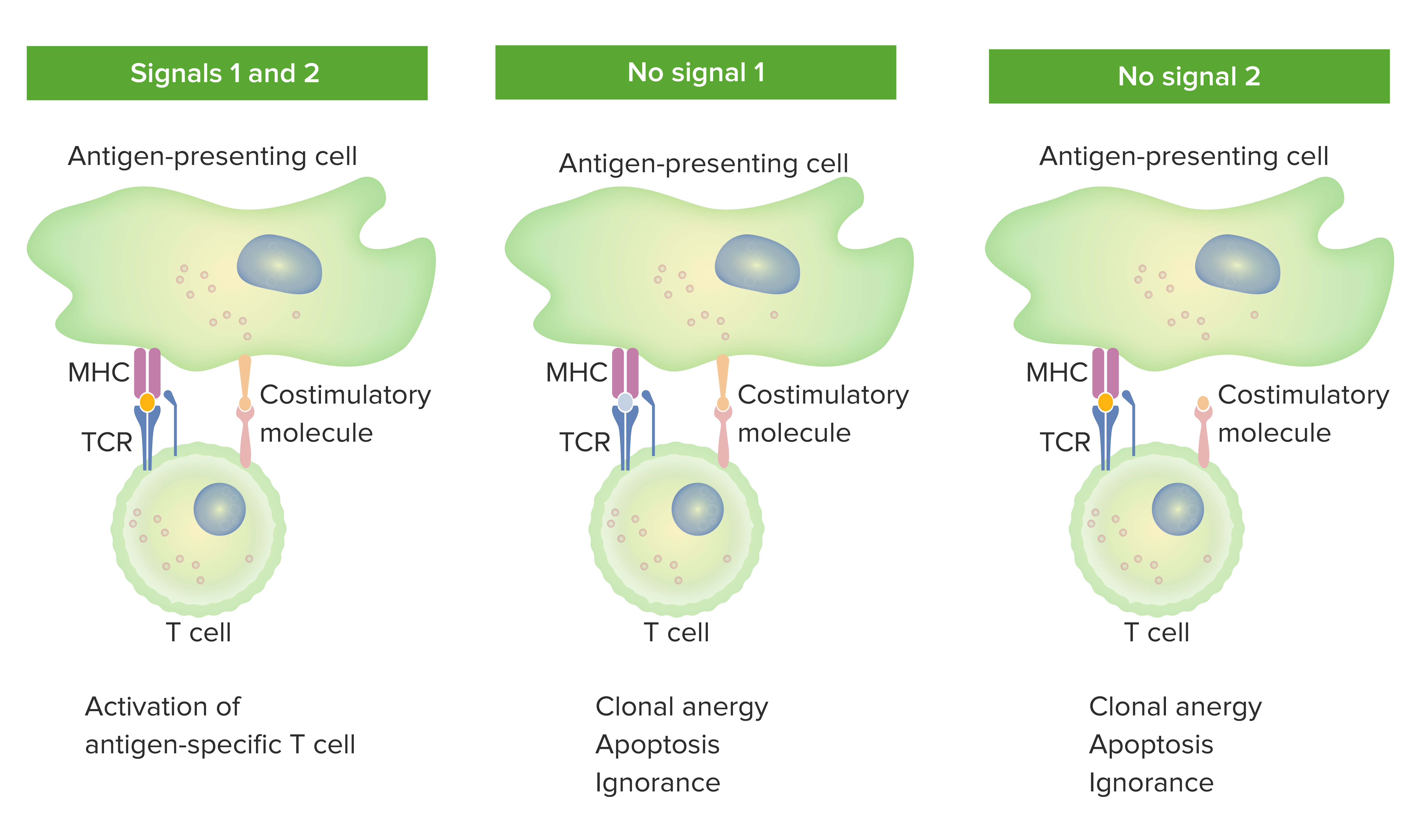
00:01 Let’s now look in a little bit more detail about the actual development process in the thymus. 00:07 We’ve heard that the T-cells migrate through the cortex and into the medulla. 00:11 So exactly what’s going on during those migration processes? Well when the T-cells first enter the thymus, having come from the bone marrow, they lack expression of two molecules - CD4 and CD8. 00:28 We therefore refer to them as being CD8 negative (CD8-), CD4 negative (CD4-). 00:34 And immunologists use a bit of jargon that’s called ‘double negative T-cell’. 00:38 So if you hear an immunologist talk about double negative T-cells, you wouldn’t know what on earth they were talking about. But what they’re actually referring to is cells that are CD4-, CD8-, lacking both of those molecules. Shortly after arrival in the thymus, the genes for both of those molecules are switched on and these T-cells become CD4 positive (CD4+), CD8 positive (CD8+); in other words in the jargon, ‘double positive T-cells’. These are now ready to undergo thymic education. 01:11 And the first step in thymic education which takes place in the cortex is interactions of the T-cells with thymic epithelial cells. 01:20 And the purpose of this interaction is to ensure that the T-cell receptor that has been generated by random recombination is able to recognize our own MHC variants. 01:34 It’s no good if the T-cell receptor can’t recognize MHC because we’re talking about alpha beta (αβ) T-cells here. 01:40 They need to recognize peptide presented by MHC. 01:43 They need to recognize peptide presented by our own variants of the MHC, not somebody else’s variant. 01:49 So the first stage in thymic education is called positive selection. 01:55 And T-cells are selected positively if they are able to recognize peptides presented by our own MHC molecules. 02:04 If they fail to do so, they die by apoptotic cell death. 02:10 So, positive selection rescues from apoptosis, cells that recognize ‘self’ MHC. 02:21 This is followed by negative selection, where there is induction of apoptosis if the T-cells recognize autoantigens, in other words, self antigens. 02:33 And this process constitutes what we refer to as central tolerance. 02:37 We say that immune cells, and we’re referring here specifically to lymphocytes; that the lymphocytes become tolerant to self antigens. 02:46 They don’t react to self antigens. 02:48 And this negative selection in the thymic medulla, where the T-cells interact with dendritic cells and macrophages that are showing self antigens to the T-cells, if there is recognition of these self antigens, apoptosis is induced and those cells are got rid of. 03:06 If the T cell had produced a T cell receptor capable of interacting with MHC class 1, then CD4 is switch off. 03:16 Whereas, if the T cell receptor is capable of interacting with MHC class 2, then CD8 is switched off. 03:23 So these T cells become what we call single positive T cells, in other words either CD4 positive or CD8 positive. 03:33 The T cells then leave the thymus and go to the secondary lymphoid tissues. 03:40 The diversity of the T cell receptor is generated by mechanisms that are essentially identical to those that generate the B cell receptor. 03:51 In other words, they are a set of T cell receptor genes that recombine. 03:58 Here we can see the numbers of gene segments that were involved. 04:07 For the T cell receptor alpha chain there are 75 variable gene segments approximately, no diversity segments, and around about 60 joining or J gene segments. 04:21 Whereas, for the beta chain, there are approximately 50 V gene segments, two diversity gene segments, and 13 J gene segments. 04:33 Regarding the gamma delta T cell receptor, there are around about 15 V segments, no Ds, 5 Js. 04:45 And for the delta chain, 8 Vs, 3 Ds and 3 Js.
About the Lecture
The lecture T-Cell Development in Thymus and Genetic Recombination of TCR – Lymphocyte Development by Peter Delves, PhD is from the course Adaptive Immune System. It contains the following chapters:
- T-Cell Development in the Thymus
- Genetic Recombination of TCR Genes
Included Quiz Questions
Which two surface molecules define ‘double positive’ T-cells?
- CD4 and CD8
- αβ T-cell receptors and γδ T-cell receptors
- Pathogen-associated molecular patterns and damage-associated molecular patterns
- Major histocompatibility complexclass I and major histocompatibility complexclass II
- CD1 and CD3
Which of the following is TRUE regarding positive and negative selection of T cells within the thymus?
- Positive selection takes place in the cortex
- Positive selection: T cells that bind "self" MHC class I or II molecules undergo apoptosis.
- Positive selection: T cells become double positive.
- Negative selection occurs in the cortex
- Negative selection: T cells that recognize and bind MHC class I or II molecules are selected.
What is the main purpose of negative selection of T cells within the thymus?
- Induction of apoptosis in thymocytes with a high affinity for "self" antigens.
- Selecting cells with a T cell receptor able to bind major histocompatibility complex class I or II molecules
- Eliminating T cells which would be non-functional due to an inability to bind major histocompatibility complex
- Eliminating thymocytes with gross defects introduced into the T cell receptor by gene rearrangement
- Silencing thymocytes with gross defects in cell surface receptors in the peripheral blood
Author of lecture T-Cell Development in Thymus and Genetic Recombination of TCR – Lymphocyte Development

Peter Delves, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
1 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
perfect and easy to follow, and friendly lecture, so far the best teacher
