Playlist
Show Playlist
Hide Playlist
Gluconeogenesis: Regulation and Glycolysis

-
05 Advanced CarbohydrateMetabolism2.pdf
-
Reference List Biochemistry.pdf
-
Download Lecture Overview
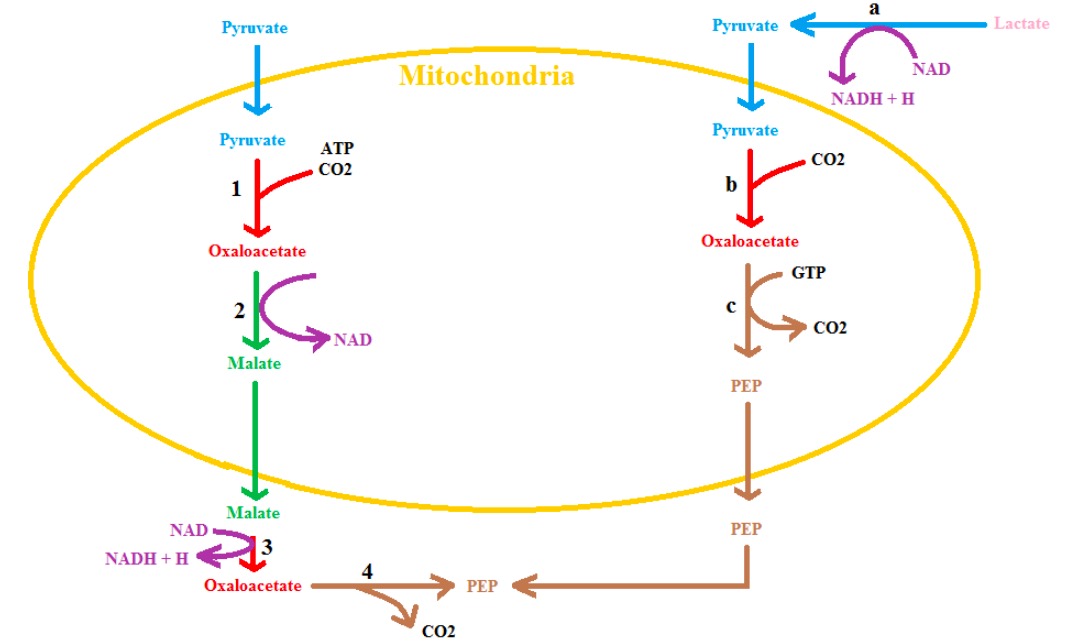
00:01 Let's turn our attention now to thinking about regulation, because, regulation is very important for making sure that these two pathways are not at loggerheads with each other. 00:11 Glycolysis goes as follows. You start with glucose 2 ADPs, 2 phosphates and 2 NADs, and you get the products shown below. 00:21 And in gluconeogenesis we are reversing the process and if you add up all the different pieces that I gave you in the pathway where we see that we start with 2 pyruvates, 2 NADHs and then it take 4 ATPs and 2 GTPs to make the glucose. 00:37 Well if you do the math what you will see that it takes more triphosphates to make glucose than you get out of glucose when you break it down. 00:46 Well that makes sense there is no process that is a 100 % efficient and this one isn't neither. 00:51 But the problem with that arises, if we turn and we try to run both these pathways at the same time. 00:57 Glycolysis makes pyruvate, pyruvate goes up to glucose, glucose goes to glycolysis and we go in this circle that's actually called a futile cycle, is what’s it called. 01:08 But this futile cycle does nothing but burn ATP and GTP. 01:15 When you burn things what happens? Heat, and loss of essential energy that you need. So the futile cycle should be avoided and that cells have interesting things setup to avoid that futile cycle. 01:30 ATP and GTP get burned as noted but nothing but heat is produced. 01:33 Now if you are trying to generate heat that could be useful. But usually you are not trying to generate heat. 01:40 So if we look at the regulatory enzymes, in glycolysis and gluconeogenesis and you are seeing the gluconeogenesis once for the first time here. 01:48 We have discovered that many of these have reciprocal effects or opposite effects on one enzyme compare to the other. 01:54 We see for example that PFK is is activated by AMP. 02:01 That makes sense. Cells need energy. They wanna get PFK going. 02:05 If cells need energy they certainly don't wanna have gluconeogenesis going because it's gonna take energy to make glucose. 02:14 Notice that FBPase gets turned off by AMP. 02:20 Notice that PEPCK gets turned off by ADP, that's a lower energy indicator than ATP. 02:25 And notice that pyruvate carboxylase is turned off by ADP. 02:31 So low energy things are turning off gluconeogenesis things and turning on glycolysis. 02:37 Hexokinase is turned off glucose-6-phosophate. That may not be apparent but the k the times when glucose-6-phosophate is high is when the cell has plenty of energy. 02:48 PFK is turned off by a high energy compound, ATP. 02:51 And pyruvate kinase, the big bang catalyzer, is also turned off by ATP. 02:59 Gluconeogenesis, on the other hand, is activated by things that indicate high energy. 03:05 Now citrate is one of these things that indicates high energy. With citrate, the fructose bisphosphatase, FBPase is activated. 03:14 Another thing that indicates high energy is acetyl-CoA. 03:19 Now acetyl-CoA you may remember is one of those things that if it's produced on x-axis can be a problem and you make fat when you have high energy and if you have a lot of acetyl-CoA, high energy. So these high energy indicators are having opposite effects on glycolysis and gluconeogenesis. 03:35 Now there is a couple of things here that don't really have any relationship or any direct relationship to energy. 03:43 The most important of these is the molecule called fructose-2,6-bisphosphate. 03:49 I mentioned in the glycolysis talk that fructose-2,6-bisphosphate was an activator of PFK. 03:55 And you will notice there is two + signs by it. It's a pretty strong activator of PFK. 04:01 You will notice also that fructose-2,6-bisphosphate has two - signs next to FBPase. 04:09 Those two enzymes catalyze equivalent reactions. 04:13 The glycolysis direction were going down from fructose-6-phosphate to fructose-1,6-bisphosphate. 04:20 In the gluconeogenesis reaction we are going up from fructose-1,6-bisphosphate back to fructose-6-phosphate. 04:26 They are catalyzing opposite things. 04:29 And notice that the regulators having opposite effects on them. 04:32 It's turning off the gluconeogenesis enzyme at the time it's turning on the glycolysis enzyme. 04:39 This reciprocal regulation is a very important concept to understand in a metabolic control. 04:46 Now one of the things about fructose-2,6-bisphosphate that I should mention is that fructose-2,6-bisphosphate is made upon insulin stimulation. 04:55 When is insulin is made? Well insulin is made when the liver for example starts getting glucose from a meal. 05:04 And you might think, "Well with the meal do I need to deal with all of that glucose?" "Should I be burning it like I am burning it here?" Well it turns out that the liver has got a little different circumstance then rest of the body. 05:14 The liver has a lot of glucose and glucose can be toxic. 05:20 So glucose must be dealt with and insulin is there to deal with glucose. 05:24 What insulin does is it stimulates the uptake of glucose and if you take up too much of glucose you will have a problem. 05:32 So cells have to deal with that access of glucose. 05:35 How do they deal with it? In two ways. One is by burning it with glycolysis and the other is by storing it in the form of glycogen. 05:45 Insulin stimulates both of those processes and result in the reduction of glucose concentration inside of cells. 05:54 Now fructose-2,6-bisphosphate is broken down by the opposite thing and the opposite this is glucagon stimulation. 06:01 Glucagon says that there is two little glucose around and the liver is gotta worry about two little glucose; because, the liver is supplying glucose to the rest of the body. 06:14 What is it doing? It favors gluconeogenesis. And what does gluconeogenesis do? It releases glucose for the body. 06:21 So the opposite effects of insulin and glucagon control the synthesis of fructose-2,6-bisphosphate and it helps supply the body with its needs exactly as needed. 06:35 In this discussion I have gone through the metabolism of other sugars as well as the synthesis of glucose. And I have talked about the ways in which these pathways can be reciprocally regulated. 06:44 By balancing the pathways, as I have described here and using very simple concepts of reciprocal regulation, the cells in our body can use and produce glucose in the way that it needs.
About the Lecture
The lecture Gluconeogenesis: Regulation and Glycolysis by Kevin Ahern, PhD is from the course Carbohydrate Metabolism.
Included Quiz Questions
Which of the following is true of futile cycles?
- Futile cycles consume energy without the net production of products.
- Futile cycles are favored when cells are running low on energy.
- Futile cycles are essential for catabolic processes.
- Futile cycles produce excess NAD+.
- Futile cycles are necessary to produce energy in the cell.
Which enzyme is activated when higher concentrations of AMP are present?
- PFK
- FBPase
- Hexokinase
- Nuclease
- Pyruvate kinase
Which enzyme is activated when higher concentrations of citrate are present?
- FBPase
- Pyruvate kinase
- Hexokinase
- Nuclease
- PEPCK
Author of lecture Gluconeogenesis: Regulation and Glycolysis

Kevin Ahern, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
2 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
A huge thank you to Prof Ahern! You explanations are very easy to understand and your lectures are a huge help to get a better understanding of biochemistry topics. Thank you very much!
Thank you Dr Ahern! Thanks to your clear and nice explanations With integrations I am finally starting to understand Biochem! You make difficult things look so simple and enhances the importance and intelligence of mother nature in our human bodies
