Playlist
Show Playlist
Hide Playlist
Cell-mediated Immunity: Gamma-Delta T-Cells, T-Cell Receptor, and Alpha-Beta T-Cells

-
Slides Cell-Mediated Immunity.pdf
-
Reference List Immune System.pdf
-
Download Lecture Overview
00:01 Let’s just spend a couple of moments to look at gamma-delta (γδ) T-cells, and exactly what they do and what they are. 00:09 They appear to have a distinct role that is complementary to that of the alpha-beta (αβ) T-cells. 00:16 They comprise less than five percent of the T-cells. 00:19 So they’re very much a minority population of T-cells. 00:22 And they’re present mostly in the gut mucosa, in the skin, in the lungs and in the uterus. 00:29 And they can directly recognize microbial pathogens, and also damaged or stressed host cells. 00:35 In other words, our own cells that are damaged due to infection or other types of pathology. 00:42 To give you just a couple of examples of the recognition that can occur by γδ T-cells. 00:48 You can have direct recognition of phosphoantigens from Mycobacterium tuberculosis, from Plasmodium malariae without any requirement for antigen processing whatsoever. 01:03 And there can be direct recognition of the MHC-like non-peptide-binding molecules, MICA and MICB, or of CD1 irrespective of any lipoprotein or glycolipid antigen. 01:17 So they can recognize structures that are coming from foreign pathogens, but they can also recognize our own molecules that are upregulated in response to stress or other events. 01:29 Here’s the structure of the T-cell receptor on the surface of a T-lymphocyte. 01:34 It’s a heterodimer, consists of two chains. 01:38 And here we can see the αβ T-cell receptor. 01:42 The γδ T-cell receptor looks almost identical. 01:49 In both cases, the two chains of the T-cell receptor are linked together by a disulfide bond. 01:56 There are transmembrane segments associated with both chains of the T-cell receptor. 02:02 Each chain is folded into two domains; a Variable domain which is stabilized by an intra-chain disulfide bond, and a Constant domain also stabilized by an intra-chain disulfide bond. 02:17 Looking at exactly how the T-cell receptor interacts with peptide MHC; at the top of this diagram, we can see the two Variable domains of the T-cell receptor. 02:31 Underneath is the MHC Class I α-chain. 02:37 This is associated with β2-microglobulin. 02:42 And then sitting between the α-helices of the MHC Class I α-chain is the antigenic peptide. 02:51 And this is making contact not only with the MHC but also with the hypervariable or CDR regions of both the α-chain and the β-chain of the T-cell receptor Variable regions. 03:06 αβ T-cells, are not all the same. There’s a variety of different types of αβ T-cells. So let us look at the different types. 03:16 Well, there are ones that have CD4 on their cell surface and there are others that have CD8 on their cell surface. 03:23 So that’s one distinction. 03:25 Within the CD4+ αβ T-cells, there are a number of subpopulations - Th1, Th2, Th17, T-follicular helper and T-regulatory cells amongst others. 03:41 These are the most important, or at least the best characterized at this point in time. 03:49 They can be distinguished by a number of different features. 03:53 So if we look at the transcription factors that are expressed in the nucleus of these different types of T-cell, we see that Th1 cells express the transcription factors T-bet and STAT4. 04:09 They produce a variety of different cytokines - gamma interferon, interleukin-2, tumor necrosis factor beta. 04:18 These are typical cytokines that are produced by Th1 cells. 04:22 And in fact, the first distinction of these different types of helper T-cells was based upon the particular cytokines that they produced. 04:31 The primary function of Th1 cells is to help cytotoxic T-lymphocytes and to help macrophages. 04:38 Although, they can also help some B-cells to produce antibody. 04:42 Importantly, they can inhibit the activity of Th2 cells. 04:48 Turning now to Th2 cells, they express the transcription factors GATA3 and STAT6 in their nucleus. 04:55 They produce the cytokines interleukin-4, IL-5, IL-6 and IL-13. 05:01 And their primary function is to help B-cells. 05:05 Importantly, just like Th1 cells can inhibit Th2 cells, so Th2 cells can inhibit Th1 cells. 05:13 So these two populations, Th1 and Th2, at least to some extent are mutually antagonistic towards each other. 05:22 Th17 cells express the transcription factor RORγt in the nucleus. 05:28 And as their name suggests, they produce interleukin-17, amongst other cytokines such as interleukin-22. 05:36 And these cells are really specialized to promote inflammation. 05:41 T-follicular helper cells are cells that are found in the germinal centers of secondary lymphoid tissues where they assist in the activation of B-lymphocytes. 05:53 They express the transcription factor Bcl-6. 05:56 And they produce cytokines such as interleukin-21 and ICOS. 06:02 And as I’ve already mentioned, their function is to help germinal center B-cells. 06:05 So those four cell types all enhance immune responses, they help immune responses. 06:12 In contrast, T-regulatory cells suppress immune responses. 06:17 And they express the transcription factor Foxp3 in their nucleus, very characteristic transcription factor of regulatory T-cells. 06:26 The cytokines they produce tend to be cytokines that overall have immunosuppressive activity. 06:34 Cytokines such as transforming growth factor beta, interleukin-10 and interleukin-35. 06:40 And again as I’ve already mentioned, the primary function of T-regs is to suppress immune responses. 06:47 So those are different types of CD4+ αβ T-cells. 06:51 What about the CD8s? Well, these are generally cytotoxic T-lymphocytes. 06:57 And they express in their nucleus, the transcription factor RUNX3. 07:03 They function by producing molecules such as perforin, granzymes, Fas ligand, but they can also produce a variety of cytokines. 07:14 And their role is to kill infected cells. 07:18 Let's take a quick look at MAIT cells or mucosal associated invariant T cells. 07:23 They can represent up to 10% of circulating T cells, but are also found in the liver. 07:28 Mucosal linings and other tissues. 07:31 They have an invariant TCR alpha chain that is linked with the restricted repertoire of TCR beta chains. 07:37 Most of these cells fall into the CD8 category and recognize antigens presented by non polymorphic MHC one like molecules MR1. 07:46 They function rapidly similar to our innate anti-microbial response, and this is mediated through the release of inflammatory cytokines listed here.
About the Lecture
The lecture Cell-mediated Immunity: Gamma-Delta T-Cells, T-Cell Receptor, and Alpha-Beta T-Cells by Peter Delves, PhD is from the course Humoral Immunity and Cell-mediated Immunity. It contains the following chapters:
- A Closer Look on Gamma-Delta T-Cells
- Structure of the T-Cell Receptor
- A Closer Look on Alpha-Beta T-Cells
Included Quiz Questions
Which of the following is a T-helper 1 specific transcription factor?
- T-bet (T-box expressed in T cells)
- Signal transducer and activator of transcription 6 (STAT6)
- RAR-related orphan receptor gamma (RORγt)
- B-cell lymphoma 6 protein (Bcl-6)
- Runt-related transcription factor 3 (RUNX3)
Gamma delta (γδ) T cells have been shown to recognize all EXCEPT which of the following?
- B-cell lymphoma 6 protein
- Mycobacterium Tuberculosis
- Plasmodium species
- Cluster of differentiation 1
- Major histocompatibility complex class I polypeptide-related sequence B
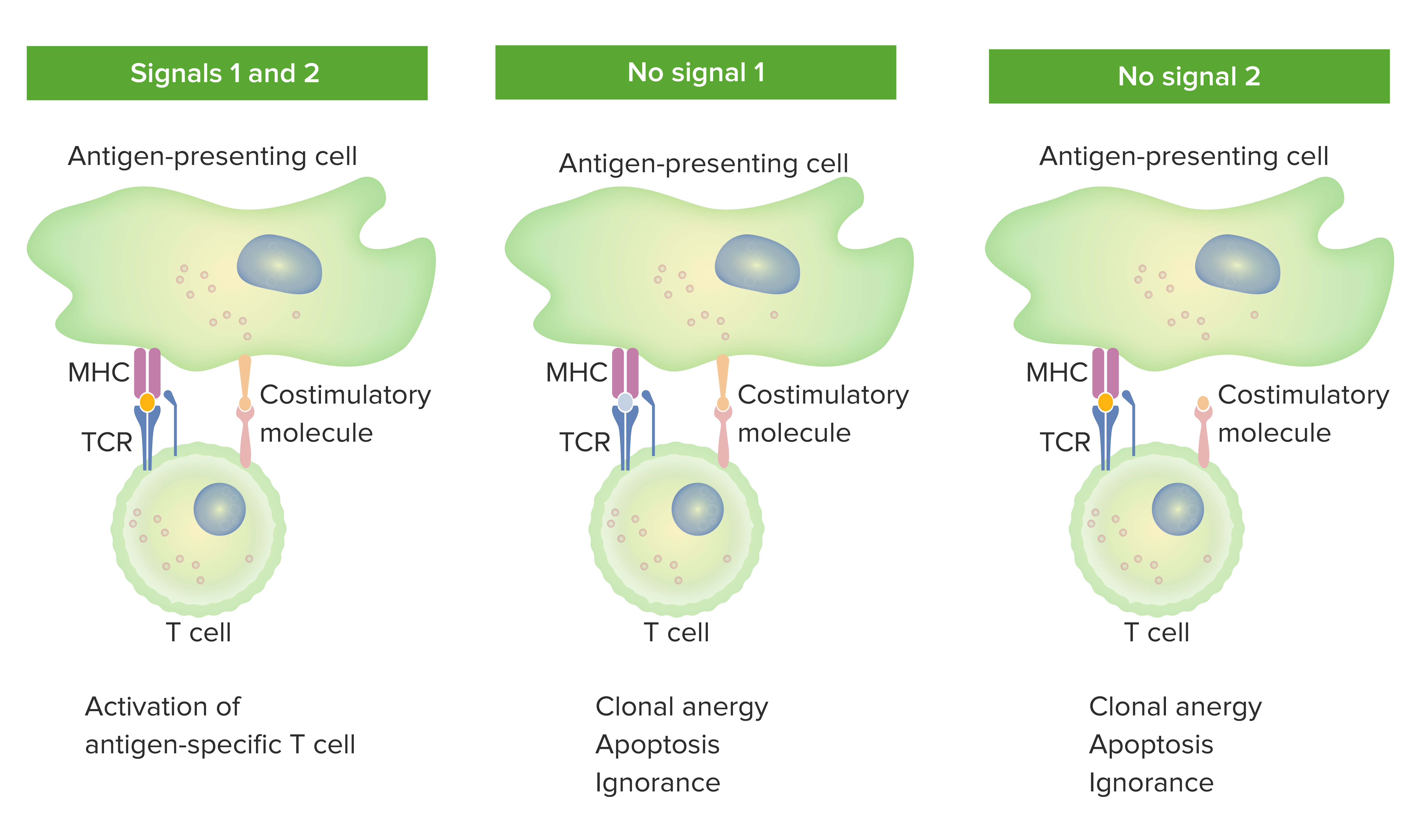
An antigen presented by a cell is connected to which part of the MHC class I molecule and the T-cell receptor, respectively?
- Antigen-binding groove, complementarity-determining regions
- Complementarity-determining regions, antigen-binding groove
- Antigen-binding groove, antigen-binding groove
- Complementarity-determining regions, complementarity-determining regions
Which of the following types of T cells dampen, rather than promote, the immune response?
- Regulatory T cells
- T helper 1
- T helper 2
- T helper 17
- Follicular helper T cells
Author of lecture Cell-mediated Immunity: Gamma-Delta T-Cells, T-Cell Receptor, and Alpha-Beta T-Cells

Peter Delves, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
1 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
Makes it way more easier to understand, it is very summarized and clear.
