Playlist
Show Playlist
Hide Playlist
Ear: Receptor Cells

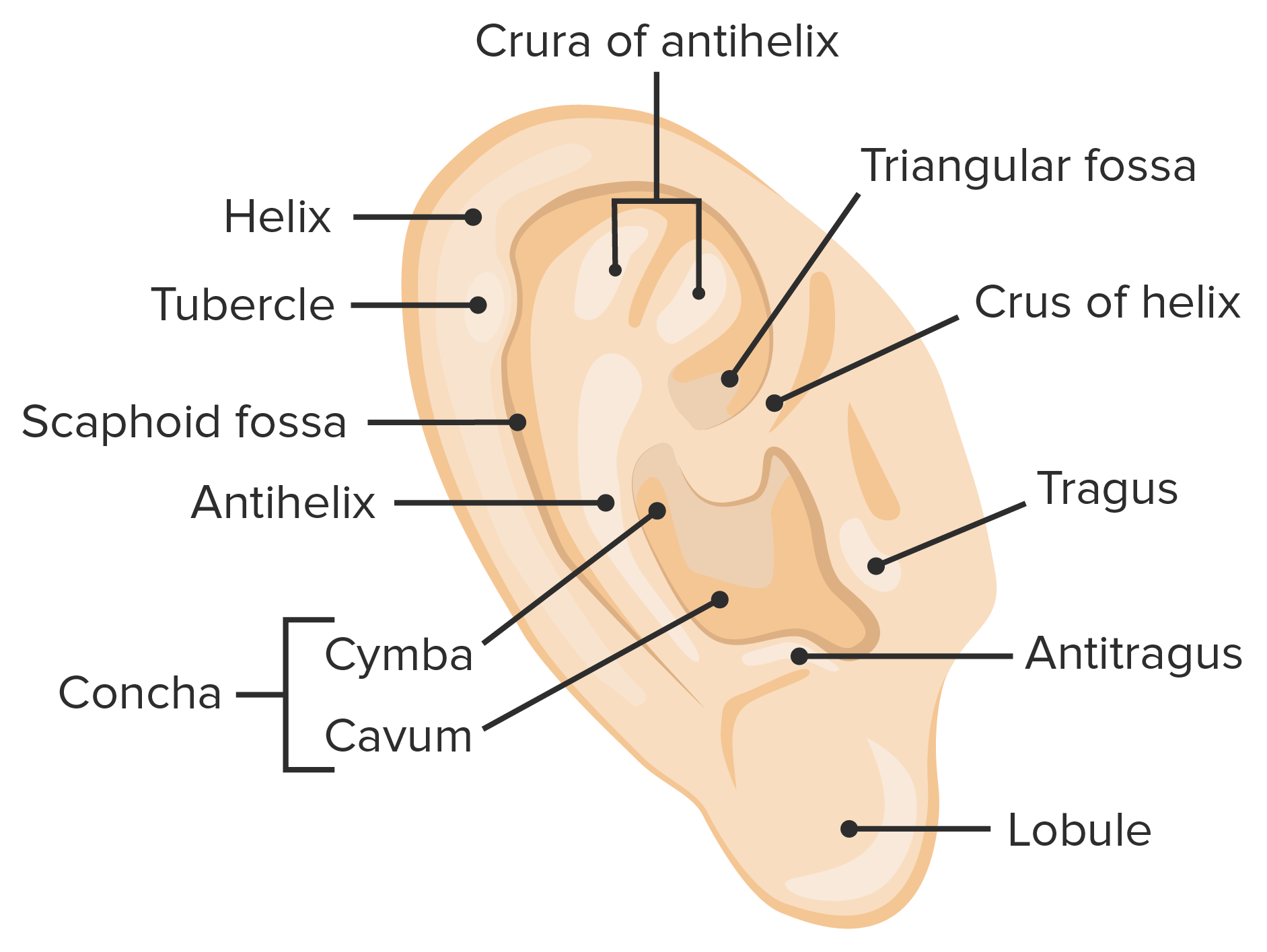
-
Slides Vestibular system.pdf
-
Reference List Histology.pdf
-
Download Lecture Overview
00:00 Before we move on and look at those sensory structures in more detail, it's important firstly to make sure we understand the histology and the ultrastructure of the hair cells. There are two hair cells shown here, type I and type II. Hair cells are very similar, whether they're in the crista ampullaris or in the maculae. They differ in the organ of Corti in one way that I'll mention at the end of this slide. But essentially, they're very similar. Type I and type II hair cells, I'm not going to really talk about any functional difference between these cells, but you can look at the slide and know that they're structurally different. 00:52 The type I hair cells have a more flask or bulbous type of appearance or shape. The type II hair cells have a more elongated shape. At their apex, they both have projections called stereocilia. And notice that these stereocilia are different in length. 01:16 They have a graded increase in length from one side of the cell to the other, and that's repeated along all the hair cells. And on the lateral, on the left lateral side of each of these hair cells, you'll see one cilium. It's called the kinocilium. That's a very important structure because that defines the polarity of the cell. 01:45 What we're going to learn later on is that those stereocilia are actually affected by movement of the endolymph when we move our head in certain directions. And that movement of the endolymph actually moves those stereocilia towards the cilium or away from the cilium. 02:07 And that sparks off opposite impulses, and it provides information about the movement of that fluid, and therefore, the position of our head. Those hair cells are supported by supporting or sustentacular cells. You can see they surround the hair cells that colored very light grey in this picture. And then the hair cells that innervate it, the bipolar neurons from the vestibular ganglion, send out a dendritic branch, the efferent nerve fibre you see here, that wrap around or form a bulbous type of connection to the type I hair cells, or simply just have isolated buttons or elongated buttons on the type II hair cells. When those hair cells are stimulated, when those stereocilia move, then those efferent fibres will send information back to the central nervous system about that movement of endolymph, and then the central nervous system interprets that into defining our position of our head in space and our movement of our head. 03:25 There is also an efferent nerve supply. These efferent fibres probably alter the sensitivity of the hair cells. They may also amend in some way the information travelling back to the central nervous system by the efferent or via the efferent nerve fibres. 03:47 The difference between these hair cells in this diagram and the hair cells that I will show you in the organ of Corti, the organ that detects sound, is that there's no kinocilium. 04:02 And that's because the stereocilia don't bend in any direction towards that kinocilium in the organ of Corti. Their impulse generation is due to hearing force and contact with the tectorial membrane that touches them on their apical surface, on the apical surface of the stereocilia. So it's a slightly different method of stimulating these hair cells. 04:31 But it does not require the kinocilium in the hair cells of the organ of Corti.
About the Lecture
The lecture Ear: Receptor Cells by Geoffrey Meyer, PhD is from the course Sensory Histology.
Included Quiz Questions
Which of the following best describes the shape of type 1 hair cells in the inner ear?
- Bulbar
- Columnar
- Flat
- Round
- Multilobular
Which of the following descriptions regarding the firing of nerve fibers at the base of the hair cells in the inner ear is MOST ACCURATE?
- Displacement of the stereocilia towards or away from the kinocilium results in changes in nerve firing.
- Displacement of the stereocilia away from the kinocilium results in increase in nerve firing.
- Displacement of the stereocilia towards the kinocilium results in decrease in nerve firing.
- Displacement of the kinocilium towards or away from the stereocilia results in changes in nerve firing.
- Displacement of the kinocilium towards the stereocilia results in decrease in nerve firing.
Stereocilia in the inner ear are affected by the movement of which of the following fluids?
- Endolymph
- Perilymph
- Air
- Blood
- Extracellular fluid
Author of lecture Ear: Receptor Cells

Geoffrey Meyer, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
5 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
