Playlist
Show Playlist
Hide Playlist
Complement System – Protein Effector Cascades

-
Slides Acute and Chronic Inflammation Effector cascades.pdf
-
Reference List Pathology.pdf
-
Download Lecture Overview
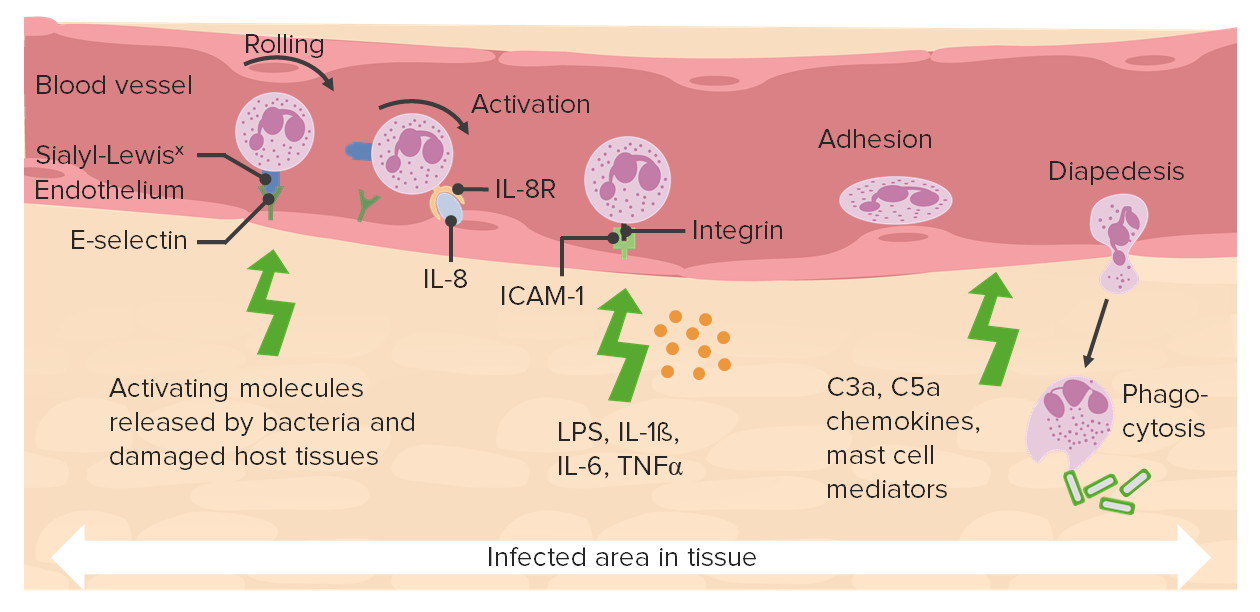
00:01 Welcome. 00:02 Previously, in this series about acute inflammation, we've talked about the vascular changes that occur. 00:08 We've talked about the cellular recruitment. 00:11 And we've talked a little bit about eicosanoids, arachidonic acid metabolites, that mediate some of the inflammatory effects. 00:19 We're going to conclude this part of our discussion of acute inflammation, looking at effector cascades. 00:27 Inflammatory cells, neutrophils don't just do their job by eating and killing, they also elaborate a variety of molecules that will drive very important downstream effects. 00:41 So acute inflammation is also about the effectors. 00:45 And that's where we're going. 00:48 There are effector cascades. 00:49 Remember that we talked about one of the truisms in biology is that you start with a little spark, and you get a cascade of ever expanding and ever increasing response. 01:03 And that's very true, the various protein effector cascades. 01:07 The proteins that we're going to be talking about come from two major sources, one is preformed and circulating in the blood. 01:14 Those are generally synthesized by the liver and excrete it into the plasma. 01:19 So there are a lot of acute phase reactants that we will revisit in other talks that are elaborated by the liver. 01:30 Then there are things that are also synthesized de novo just at the site of action. 01:36 So many of the cells directly in the line of fire, at a site of injury will elaborate proteins that also have an effector cascade. 01:49 The major cell that's synthesizing them, yet that site are going to be the macrophages. 01:54 So we have preformed circulating mediators from the liver macrophages synthesizing things as we need them, where we need them. 02:03 These proteins from both sources include complement, coagulation factors, and kinins. 02:11 And those are the three cascades that we're going to be talking about now. 02:16 Notably, they're synthesized by the liver, and by macrophages. 02:21 So we're talking about the same thing in different places. 02:26 Let's talk first about complement-activation. 02:28 There are many different ways to activate the complement cascade. 02:33 In the next few slides, we'll talk about how we do that and then exactly what that cascade involves. 02:38 There is a classical pathway, where antibody indicated here is the orange bivalent molecules binding to an antigen on the surface of the microbe will undergo a conformational change that will allow the activation of complement will start that cascade. 02:57 And complement activation is basically a variety, a sequence of proteases proteolytic cleavages to give us activated fragments. 03:06 The key linchpin is going to be C3 fragmentation C3 proteolysis, to give a C3a and C3b. 03:17 And we'll come back to that. 03:18 So the classical pathway involves antibody binding, and then complement being activated because of an antibody binding. 03:25 There's another pathway called the alternative pathway to be compared with the classical pathway. 03:31 And this involves additional proteins called factor B, factor D, and properdin. 03:39 That will also get us to that C3b proteolysis, but by a slightly different set of factors. 03:46 And then there's the lectin pathway. 03:48 And this has to do with mannose-binding lectin. 03:52 So recall that many of the proteins, glycoproteins on the surface of pathogens, and in a terminal mannose. 04:01 We know that, and our liver and macrophages synthesize a mannose-binding protein or mannose-binding lectin that will then act as a trigger once it's bound to a pathogen to activate the complement. 04:14 Again, a proteolytic cleavage from C3 to C3b and C3a. 04:19 C3a and C3b are just fragments of the original C3 molecule. 04:25 Okay, that's the way that we got it activated. 04:27 Now what happens? Well, so C3a and C3b so that activated C3b on the surface will drive the subsequent breakdown proteolysis, and a cascade of additional complement fragments including C5. 04:45 C3a and C5a are very potent chemotactic agents. 04:51 They will recruit and activate neutrophils, leukocytes that will lead to the destruction by those leukocyte destruction of the microbes by those leukocytes. 05:01 So it's pro inflammatory C3a and C5a. 05:05 C3b, once it gets generated is very sticky, and on the surface of a microbe acts as an obstinate, it makes it tasty. 05:17 So now, cells that have receptors for C3b such as neutrophils, and macrophages can bind, and will ingest will phagocytize the microbe and kill it. 05:29 And finally, starting with C3b cleavage of C3, to C3a and b. 05:35 Starting with that, we will then cleave C5, and C6, and C7, and C8 and that will form a membrane attack complex. 05:44 So we get a pore and that MAC or Membrane Attack Complex will lead to lysis of microbes. 05:50 So complement does a variety of things. 05:52 It's pro inflammatory by recruiting inflammatory cells. 05:55 It makes microbes tasty, so that we can ingest them. 05:59 And it will also directly lyse microbes. 06:04 So very important. 06:07 So complement. 06:08 Promised that we will give an overview of this. 06:10 It's a collection of circulating and macrophage-synthesize proteins. 06:15 It's a cascade of activation triggered by Antigen-antibody complexes, bacterial polysaccharides. 06:22 that's that mannose-binding protein, and aggregated IGA. 06:26 A form of immunoglobulin. 06:28 So that involves the properdin alternate pathway. 06:32 Okay. 06:33 Once they're activated, they have a variety of effects. 06:36 They will cause vasodilation increased permeability. 06:40 They will increase leukocyte adhesion. 06:42 Promote chemotaxis and activation of leukocytes. 06:45 And they will induce opsonization. 06:49 Okay, so complement does a lot of stuff.
About the Lecture
The lecture Complement System – Protein Effector Cascades by Richard Mitchell, MD, PhD is from the course Acute and Chronic Inflammation.
Included Quiz Questions
Which of the following activates the classical pathway of the complement cascade?
- Antigen-antibody complexes
- Factor B
- Factor D
- Properdin
- Mannose-6-phosphate
Which of the following complement fragments facilitates phagocytosis?
- C3b
- C5a
- C5b
- C3a
- C4a
Which of the following induces proinflammatory responses in the tissues?
- C5a
- C5b
- C4a
- C3b
- C5b-6-7-8 complex
Where are complement proteins synthesized?
- Liver
- Spleen
- Kidneys
- Lymph nodes
- Pancreas
Author of lecture Complement System – Protein Effector Cascades

Richard Mitchell, MD, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
5 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
