Playlist
Show Playlist
Hide Playlist
Receptors and Messengers

-
Slides HormonesSignalTransduction Biochemistry.pdf
-
Reference List Biochemistry.pdf
-
Download Lecture Overview
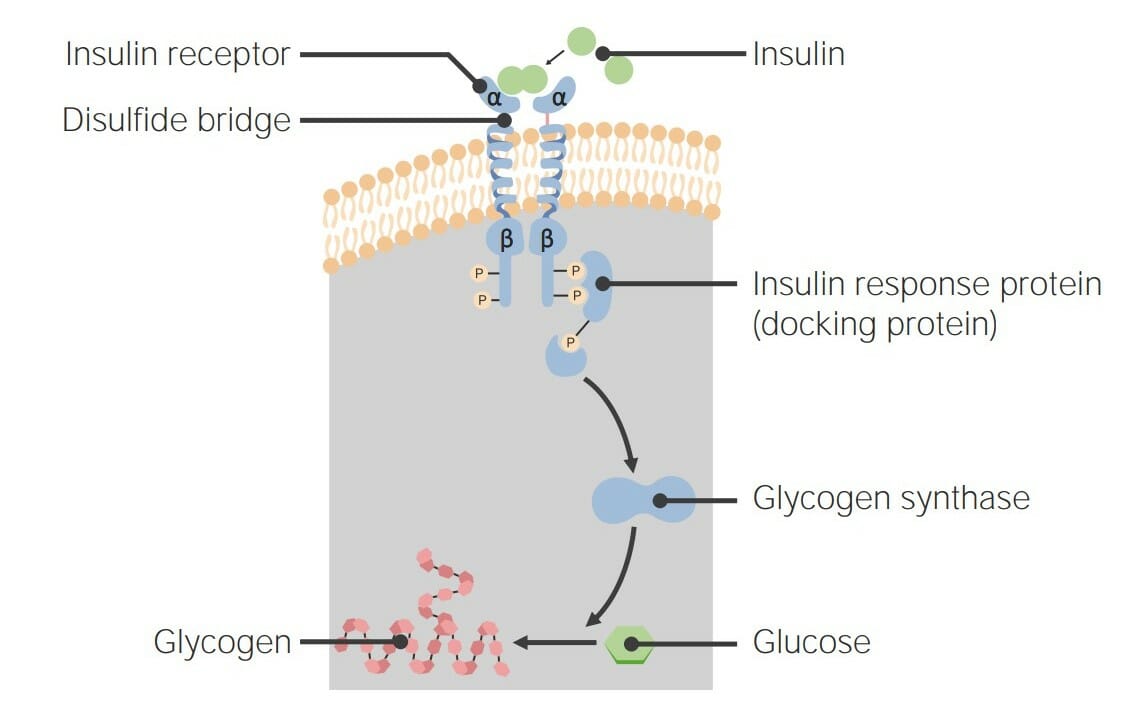
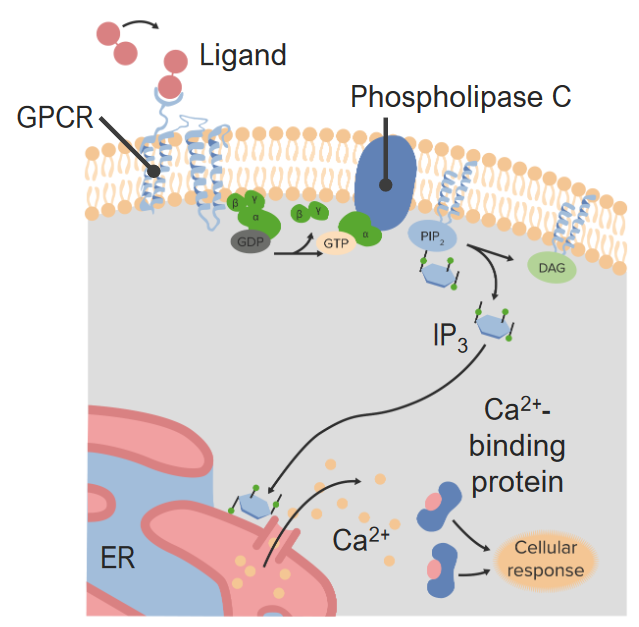
00:01 Dieses Bild zeigt einen Hormonrezeptor in orange und ein Hormon in gelb, das an ihn bindet. 00:07 Wie ich vorher bereits sagte, sind die meisten Hormonrezeptoren membrangebunden, eingebettet in die Membran. 00:12 Und die meisten Steroidhormonrezeptoren befinden sich im Zytoplasma oder im Zellkern. 00:17 Es ist die Wechselwirkung des Hormons mit dem Rezeptor, die das Signal auslöst, über das das Hormon letztlich kommuniziert und innerhalb der Zelle ausgeübt werden soll. 00:26 Diese einfache Abbildung zeigt schematisch, was hier passiert. 00:30 Auf der linken Seite sehen wir, dass das Hormon sich bereit macht, an den Rezeptor zu binden. 00:35 Und auf der rechten Seite sehen wir das Hormon, nachdem es an den Rezeptor gebunden hat. 00:39 Auf der Außenseite der Zelle, die hier die obere Schicht ist, das ist wo das Hormon gefunden wird, sehen wir, dass es eine Bindungsstelle für dieses Hormon gibt. 00:47 Und auf der Unterseite des Rezeptors, das ist die Innenseite der Zelle, sehen wir, dass nach der Bindung des Hormons dieser Teil des Proteins seine Form sehr leicht verändert. 00:58 Eine sehr geringe Veränderung der Form dieses Proteins verändert die Interaktion des Proteins mit Molekülen, die sich im Inneren der Zelle befinden. 01:07 Was wir hier sehen, ist also der erste Schritt der Kommunikation und wir sehen, wie diese Kommunikation tatsächlich letztendlich ihre Wirkung entfaltet. 01:16 Dieses Bild zeigt also zwei verschiedene Wege, auf denen Signale innerhalb der Zelle kommuniziert werden können. 01:23 Auf der linken Seite sehen wir Steroidhormone und wie ich schon sagte sind Steroidhormone ungewöhnlich, da sie in der Lage sind, sich selbstständig durch diese Lipiddoppelschicht zu navigieren. 01:32 Sie gelangen in das Zytoplasma, das diese mittlere Schicht ist, die sich dort befindet. 01:36 Und Sie sehen, dass sie sich an das lila gefärbte Molekül binden. 01:39 Das lila gefärbte Molekül ist ein Steroid- rezeptor, der im Zytoplasma herumschwimmt. 01:46 Wenn der Steroidrezeptor an das Hormon bindet, können Sie sehen, dass er sich nach unten bewegt und nach unten bewegt er sich in Richtung des Zellkerns, denn Steroidhormon- rezeptoren gehen und binden direkt an die DNA und stimulieren die Expression von spezifischen Genen, was dazu führt, dass die Aktionen der Hormone schlussendlich ausgeführt werden. 02:06 Die andere Abbildung, die auf dem Bildschirm angezeigt wird, zeigt, was mit einem Rezeptor passiert, der kein Steroidhormonrezeptor ist. 02:13 Hier sehen wir, dass es sich um einen membrangebundenen Rezeptor handelt. 02:17 Zur Orientierung: Der äußere Teil der Zelle ist nach oben gerichtet und wie zuvor, bewegen wir uns nach unten, nähern wir uns dem Kern. 02:24 Sie können das Hormon sehen, das in der Lösung schwimmt und Sie sehen ein Hormon, das an seinen Rezeptor gebunden hat. 02:30 Darunter sehen wir auch eine Reihe von verschiedenen Kästchen und Kreisen und Farben und so weiter; jedes einzelne dieser Moleküle ist ein eigenes Protein. 02:41 Nun hat die Bindung der Membran an den Rezeptor zur Bildung eines Komplexes geführt. 02:47 Und dieser Komplex ist beteiligt daran, ein Signal nach unten zu übermitteln. 02:52 Wenn wir also diesen membrangebundenen Rezeptor betrachten, sehen wir, dass die Komplexität des Signals, das sich durch die Zelle bewegt, sehr hoch sein kann. 03:00 Wenn wir beispielsweise darüber nachdenken, wie sich unsere Computer mit dem Internet verbinden, denken wir, es muss zum Beispiel über WiFi geschehen und dieses WiFi muss mit einem anderen System verbunden werden und dieses wiederum mit einem anderen System usw. 03:10 Jede dieser Verbindungen ist ein Knotenpunkt in der Gesamtverbindung Ihres Computers mit einem größeren Computer anderswo. 03:16 Nun, das ist sehr ähnlich zu dem, was hier passiert, aber überraschenderweise gibt es eine ganze Reihe von Proteinen, die an diesem Prozess beteiligt sind und wir werden sehen, wie das abläuft. 03:25 Nachdem also der erste Rezeptor an das Hormon gebunden hat, muss das Signal nach innen weitergeleitet werden. 03:32 Und wie Sie in dem von mir gezeigten Beispiel für ein Protein, das ein Membranrezeptor war, gesehen haben, gab es andere Proteine auf dem Weg, die Signale übermittelten. 03:40 Ich möchte Sie auch daran erinnern, dass nicht alle der bei der Übermittlung von Nachrichten beteiligten Moleküle Proteine im Inneren der Zelle sind. 03:48 Einige der Kommunikatoren werden zweite Boten genannt. 03:52 Und diese Botenstoffe sind kleine Moleküle, die für die Übermittlung der Informationen zuständig sind. 03:58 Einige von ihnen können wir hier sehen. 04:01 Zuerst sehen wir eines mit dem Namen IP3. 04:03 Es ist relativ klein. 04:04 Wenn ich klein sage, dann meine ich kleiner als ein Eiweiß. 04:08 Ein weiterer, sehr häufig vorkommender Stoff innerhalb von Zellen ist zyklisches AMP. 04:12 Es ist verwandt mit dem Molekül AMP und dem Molekül ATP, hat aber eine zyklische Gruppe, wie Sie auf der linken Seite des Moleküls sehen können. 04:21 Kalzium, wie ich in anderen Vorlesungen erwähnt habe, erfüllt ebenfalls Funktionen der zweiten Botenstoffe und zwar durch die Freisetzung in das Zytoplasma der Zelle. 04:31 Ein weiteres zyklisches Nukleotid, das an der Übermittlung von Informationen beteiligt ist, ist zyklisches GMP. 04:37 Und wir haben in einem anderen Vortrag darüber gesprochen, wie zyklisches GMP bei der Übermittlung von Informationen im Prozess des Sehens hilft. 04:46 Die kovalente Modifikation findet also während der Übermittlung der Nachricht statt. 04:51 Auf der vorherigen Folie habe ich eine eine Vielzahl von Proteinen gezeigt, die vom einen zum anderen, zum anderen, zum anderen, zum anderen und schließlich zum Kern gehen. 05:00 Was passiert hier? Nun, was passiert, ist, dass jedes dieser Proteine chemisch verändert wird, während diese Information übermittelt wird. 05:09 Diese chemische Modifikation ist normalerweise ein Phosphat, das entweder auf ein Protein aufgebracht oder von ihm entfernt wird. 05:16 Und diese chemische Veränderung ist im Wesentlichen das Signal. 05:20 Das Signal ist also, wie Sie wahrscheinlich zu erkennen beginnen, komplex. 05:24 Es kann in Form von Botenstoffen kommen, den kleinen Molekülen, die Sie hier sehen. 05:28 Es kann auch in Form von chemischen Modifikationen kommen, die auch an Proteinen vorgenommen werden. 05:34 Nun, eines der Dinge, die schlussendlich in der Kommunikation von vielen Nachrichten passieren, das ist die Veränderung des Musters der Genexpression in einer bestimmten Zelle. 05:44 Erinnere dich daran, dass eine Zelle in einem mehrzelligen Organismus verschiedene Gene hat, die sie zu verschiedenen Zeiten exprimiert. 05:50 Eine Knochenzelle zum Beispiel exprimiert andere Gene, als eine Muskelzelle. 05:55 Kontrollieren zu können, welche Gene exprimiert werden, erlaubt der Zelle also entsprechend der Bedürfnisse des Körpers und der individuellen Bedürfnisse der Zelle zu reagieren. 06:06 Ein weiterer Weg, auf dem Hormone kommunizieren und den Weg der Zelle verändern können, ist durch Veränderung der Aktivität der Enzyme in der Zelle und wir werden gleich ein Beispiel dazu sehen, wie der Prozess des Glykogenstoffwechsels durch die Einwirkung eines Hormons drastisch verändert werden. 06:26 Diese Abbildung zeigt also ein einfaches Schema von zweiten Botenstoffen (Second Messengers), in Verbindung mit einem Protein namens Phospholipase C. 06:34 Phospholipase C ist ein Enzym, das in der Zellmembran zu finden ist und durch Hormoneinwirkung aktiviert wird. 06:41 Dies ist ein sehr schnell wirkender Prozess, denn bei diesem Vorgang wird ein Enzym aktiviert, die Phospholipase C. 06:49 Wenn sie aktiviert ist, katalysiert die Phospholipase C eine Reaktion auf ein Molekül namens PIP2 in der Lipiddoppelschicht der Zelle. 06:58 Diese von der Phospholipase C katalysierte Reaktion spaltet PIP2 in zwei Moleküle. 07:05 Eines der Moleküle ist als IP3 bekannt, und Sie können sehen, wie es das Zytoplasma der Zelle durchquert und sich nach unten in Richtung des endoplasmatischen Retikulums, eine Organelle, die zum Beispiel Kalzium speichert, begibt. 07:18 Wenn IP3 zu einem Rezeptor auf der Oberfläche des endoplasmatischen Retikulums gelangt, bindet es daran und bewirkt die Freisetzung von Kalzium in das Zytoplasma der Zelle. 07:30 Kalzium ist auch ein zweiter Botenstoff. 07:32 Wir haben also gesehen, dass es jetzt zwei zweite Botenstoffe gibt, einer ist IP3 und der Zweite ist das Kalzium, das vom endoplasmatischen Retikulum freigesetzt wird. 07:40 Das Kalzium geht und bindet an ein Protein, das als Proteinkinase C bekannt ist. 07:46 Die Proteinkinase C ist ein Protein, das andere Proteine physikalisch verändern kann. 07:52 Erinnern Sie sich, daß ich sagte, wir können andere Proteine kovalent verändern. 07:55 Das ist also was passiert, wenn Kalzium an die Proteinkinase C bindet. 08:00 Nun, damit die Proteinkinase C voll aktiv sein kann, braucht sie einen zusätzlichen zweiten Botenstoff, der ein dritter zweiter Botenstoff ist. 08:08 Und der dritte Botenstoff, der verwendet wird, ist in die Lipiddoppelschicht der Zelle eingebettet und als DAG bekannt. 08:16 Nun war DAG nicht zufällig das Produkt der Katalyse von Phospholipase C an PIP2. 08:24 Wenn Phospholipase C also PIP2 spaltet, zerlegt sie es in IP3 und DAG. 08:30 Beide gingen an unterschiedliche Orte, um ihre Wirkung zu entfalten. 08:34 Aber der koordinierte Zweck war die Aktivierung des als Proteinkinase C bekannten Proteins. 08:40 Die Proteinkinase C kann andere Proteine phosphorylieren und die Zelle veranlassen, ihre Genexpression zu verändern. 08:48 Dieses Muster der Modifizierung von Proteinen kann sehr komplex sein. 08:52 Es ist erstaunlich, welches Netzwerk von individuellen Prozessen im Inneren der Zellen abläuft. 08:57 Also keine Sorge, ich werde Sie nicht durch all das führen. 09:00 Es genügt zu sagen, dass wir wahrscheinlich einen ganzen Kurs über das, was auf dieser einen Folie hier gezeigt wird, abhalten könnten. 09:05 Aber die Botschaft, die Sie mitnehmen sollen ist, dass es in diesem Prozess, den Sie hier sehen, mehrere Komponenten gibt, die alle interagieren und eine Bemühung koordinieren, die sich in diesem Fall auf den Kern konzentriert. 09:18 Und der Zellkern ist natürlich der Ort, an dem die DNA gespeichert ist, und und dort wird die Genexpression als Ergebnis der Prozesse, die durch diese Hormonsignalprozesse ablaufen, gesteuert. 09:28 Die zelluläre Signalübertragung ist sehr komplex. 09:32 Und die Antworten sind in jedem Fall darauf ausgerichtet, dem Organismus zu nützen, genau wie das Militär wo ich einen einzelnen Soldaten habe. 09:41 Der Soldat bemüht sich, der Einheit als Ganzes zu helfen. 09:45 Auch die Aktionen der Zellen sind darauf ausgerichtet, den Organismus als Ganzes zu unterstützen.
About the Lecture
The lecture Receptors and Messengers by Kevin Ahern, PhD is from the course Hormones and Signal Transduction. It contains the following chapters:
- Receptors
- Receipt of Message
- Second Messengers
Included Quiz Questions
Which statement regarding hormone receptors is true?
- They change shape when binding occurs.
- They are always membrane bound.
- They change shape on binding to another receptor.
- They require co-factors to be stimulated.
- They are specific to a single type of substance.
Which statement regarding second messengers is true?
- They use covalent interactions to modify proteins.
- They use hydrogen interactions to modify proteins.
- They carry messages out of cells.
- They use van der Waals forces to modify proteins.
What acts as a second messenger inside the cell? Select all that apply.
- Inositol-1, 4, 5-triphosphate
- Deoxyribose nucleic acid
- cAMP
- cGMP
- Ca+2 ions
What kind of modification occurs during the communication of signals via the second messenger molecules in the case of non-steroid hormones?
- Covalent changes in protein molecules.
- Hydration of protein molecules.
- Dehydration of protein molecules.
- Conversion of triple bonds to double bonds between the 2 carbon atoms of a protein molecule.
- Conversion of triple bonds to single bonds between the 2 carbon atoms of a protein molecule.
Which statement regarding phospholipase C is true?
- An activated phospholipase C enzyme catalyzes the breakdown of the PIP2 molecule to IP3 and DAG.
- Phospholipase C protein acts as a second messenger that directly stimulates the gene expressions by binding with the DNA molecules.
- Phospholipase C protein acts as a second messenger molecule for the synthesis of PIP2 from DAG and IP3.
- Phospholipase C acts as an energy source for the transduction of extracellular response toward the Golgi apparatus.
Author of lecture Receptors and Messengers

Kevin Ahern, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
1 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
Thank you Dr. Kevin Ahern!!! Sometimes you speak so fast, but these time was perfect :)
