Playlist
Show Playlist
Hide Playlist
Hormonal Control

-
11 Advanced AdvancedMetabolicControl.pdf
-
Reference List Biochemistry.pdf
-
Download Lecture Overview
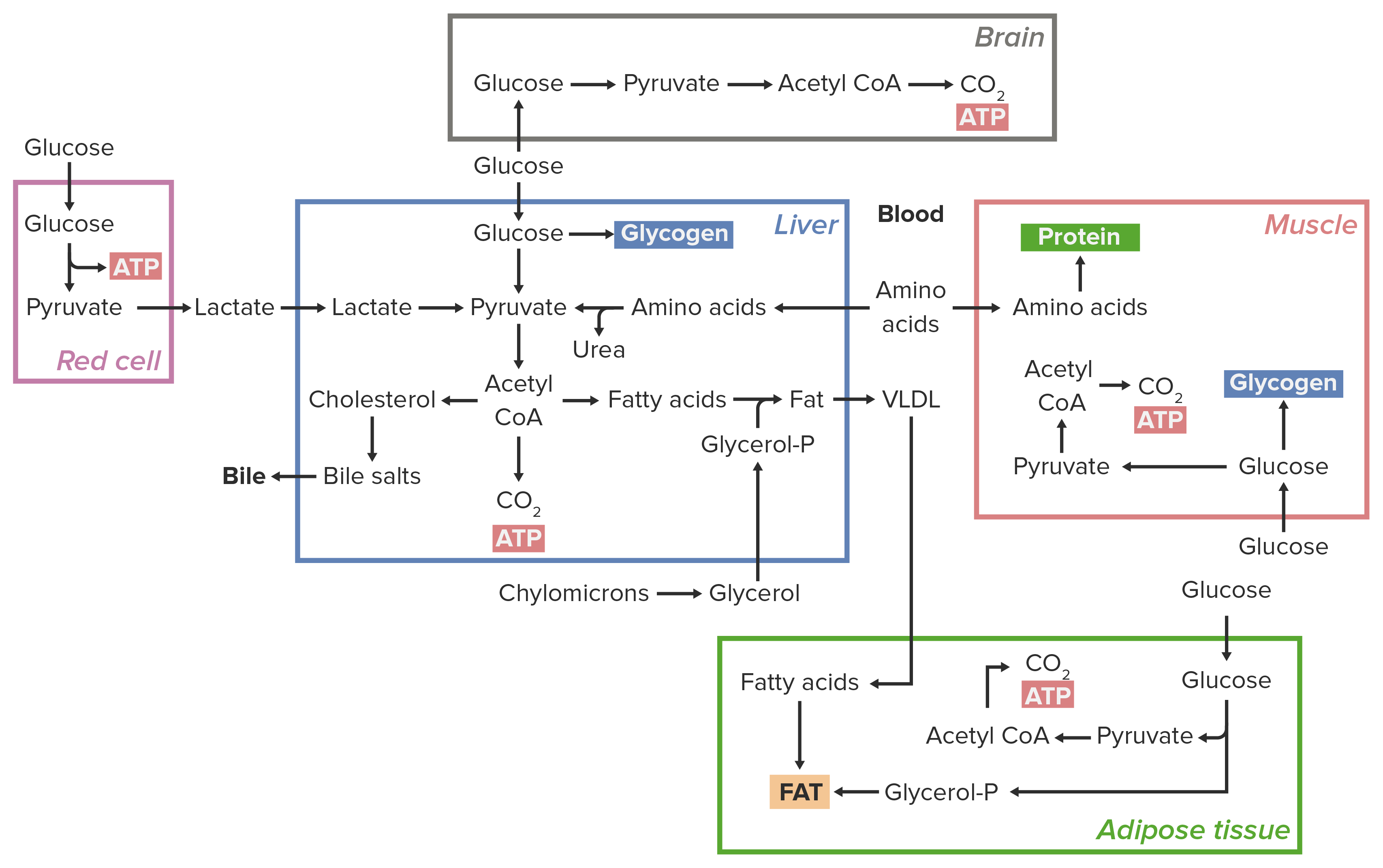
00:01 Jetzt möchte ich über zwei Organe sprechen, die eine wichtige Rolle bei der hormonellen Steuerung spielen, über die wir hier sprechen wollen. 00:08 Dies sind die Nebennieren, in denen das Adrenalin synthetisiert wird. Epinephrin ist ein Hormon, das auch als Adrenalin bekannt ist. 00:17 Es wird synthetisiert, wenn der Körper versucht, eine Menge Glukose zu produzieren. 00:22 Insulin ist eine Art Gegenhormon zu Epinephrin. 00:25 Insulin wird produziert, wenn der Körper versucht, die Konzentration von Glukose im Blut zu senken und es wird in der Bauchspeicheldrüse synthetisiert. 00:35 Es ist nun sehr wichtig, dass der Körper einen relativ konstanten Glukosespiegel aufrechterhält. 00:42 Glukose wirkt im Körper wie ein Gift. Zu viel davon ist nicht gut. 00:48 Wenn also der Blutzuckerspiegel zu hoch wird, dann führt dies zu Problemen. Deshalb gibt es Insulin. 00:55 Ein zu niedriger Wert ist aber auch ein Problem, weil man hypoglykämisch werden kann. 01:00 Deshalb sind Hormone wie das Adrenalin und das Glukagon, die die Produktion von Glukose stimulieren, in einem mehrzelligen Organismus für dieses Gleichgewicht unerlässlich. 01:09 Die hormonelle Kontrolle, die wir sehen werden, wird also weitgehend durch die unterschiedlichen Aktivitäten dieser beiden Hormone und deren Auswirkungen auf die Enzyme im Stoffwechsel bewirkt. 01:20 In dem Schema der Glykolyse und Glukoneogenese, das ich vorhin gezeigt habe, ist das Molekül Fructose-2.6-Biphosphat oder F2.6BP, das einen wichtigen wechselseitigen Regulator darstellt. 01:32 Wie sich herausstellt, ist dieses Molekül fast der wichtigste wechselseitige Regulator der beiden Wege. 01:38 Es ist also wichtig für uns zu verstehen, wie es das tut, was es tut und wie es hergestellt und abgebaut wird. 01:45 F2.6BP ist also ein Aktivator der PFK, dem wichtigsten regulatorischen Enzym der Glykolyse. 01:53 Es ist ein Hemmstoff der Fructose-1,6-Bisphosphatase, dem entsprechenden Enzym der Gluconeogenese. 02:01 Es ist also wichtig zu verstehen, was mit dem Fructose-2,6-Bisphosphat passiert, denn wenn man es bildet, wird der eine Weg bevorzugt und wenn man es abbaut, wird der andere Weg bevorzugt. 02:11 Dieses Molekül wird von einem sehr interessanten Enzym hergestellt. Es wird von einem Enzym hergestellt, das zwei Aktivitäten hat. 02:20 Es wird also von demselben Enzym hergestellt und abgebaut. Die beiden Aktivitäten arbeiten zusammen. 02:27 Dies zeigt die Synthese von Fructose-2.6-Bisphosphat aus Fruktose-6-Phosphat. 02:33 Dies ist nun keine Reaktion der Glykolyse, obwohl Fruktose-6-Phosphat ein Zwischenprodukt der Glykolyse darstellt. 02:41 Die Reaktion wird durch das Enzym katalysiert, das Sie hier sehen und das ich mit einem Yin und Yang gezeichnet habe, weil das Enzym sich so verhält. 02:49 Sie werden feststellen, dass oben ein blauer Teil zu sehen ist, wo PFK 2 steht. 02:53 Die PFK 2 katalysiert also die Addition von einem Phosphat an Position 2 an das Fructose-6-Phosphat, um Fructose-2.6-Bisphosphat herzustellen. 03:05 Fruktose-2,6-Bisphosphat wird durch den gelben Teil des Enzyms namens FBPase 2 abgebaut. 03:15 Wenn Sie sich diese beiden Enzyme genau ansehen, werden Sie sehen, dass sie unterschiedlich sind. 03:19 Der Unterschied ist, dass sie ein P in diesem kleinen Kreisen haben. Dieses P bezieht sich auf ein Phosphat. 03:26 In diesem Fall bestimmt die Zugabe eines Phosphats zu diesem Enzym, welche der beiden Aktivitäten tatsächlich abläuft. 03:34 Wenn kein Phosphat vorhanden ist, ist die PFK 2 aktiv. Wenn das Phosphat vorhanden ist, ist die Fructosebisphosphatase 2, das Enzym, das die Reaktion linker Hand katalysiert, aktiv. 03:48 Diese Folie zeigt, wie das Phosphat an das Yin-Yang-Enzym angelagert und von diesem Yin-und-Yang-Enzym abgezogen wird. 03:55 Es gibt ein Protein namens Proteinkinase A, das das Phosphat an das Yin-und-Yang-Enzym weitergibt. 04:01 Der Weg, auf dem das Phosphat angefügt wird, wird durch das Hormon Epinephrin oder das Hormon Glucagon aktiviert. 04:08 Sie wirken auf sehr ähnliche Art und Weise. Beide versuchen abwechselnd, die Konzentration von Glukose im Körper zu erhöhen. 04:16 Das Phosphat des Enzyms FBPase 2 wird von einem Protein namens Phosphoproteinphosphatase entfernt. 04:24 Und wenn Sie darüber nachgedacht haben, was ich vorhin gesagt habe, werden Sie wahrscheinlich bemerken, dass es durch Insulinstimulation aktiviert wird. 04:30 Epinephrin wird also die Phosphorylierung begünstigen und Insulin die Dephosphorylierung. 04:37 Wir werden sehen, dass es ein gemeinsames Muster gibt in einigen der Pfade, über die ich hier sprechen werde. 04:41 Wenn Epinephrin oder Glucagon an eine Leberzelle binden, wird Fruktose-2,6-Bisphosphat abgebaut. Unter diesen Bedingungen wird die Glykolyse, wie wir bereits bei der Regulierung gesehen haben, gehemmt. 04:54 Wenn nun die Glykolyse gehemmt ist, wird die Gluconeogenese begünstigt. Bedenken Sie hierbei, dass Epinephrin und Glucagon produziert werden um den Glukosespiegel des Körpers anzuheben. 05:06 Wenn Insulin an eine Leberzelle bindet, wird Fructose-2,6-Bisphosphat gebildet. 05:13 Unter diesen Bedingungen wird die PFK stimuliert, sodass die Glykolyse begünstigt wird und das Insulin dazu führt, dass der Glukosespiegel im Körper gesenkt wird. Die Verbrennung durch Glykolyse macht also sehr viel Sinn. 05:26 Wir haben in anderen Präsentationen über den Weg gesprochen, über den die Hormonstimulation außerhalb der Zelle die Informationen innerhalb der Zellen weitergibt. 05:34 Jetzt werde ich das Ganze noch einmal darauf zurückführen, um zu sehen wie sich das auf die Enzyme auswirkt, über die wir gerade gesprochen haben. 05:39 In diesem Fall handelt es sich um eine Leberzelle, an deren Rezeptor Epinephrin oder Glucagon gebunden hat. 05:46 Wenn das am Rezeptor passiert, wird das G-Protein, das an den Rezeptor gebunden ist, aktiviert. 05:52 Es wird aktiviert, indem es ein paar seiner Untereinheiten loslässt und sein GDP durch GTP ersetzt. 05:59 Wenn das passiert, stimuliert das G-Protein ein Enzym in der Membran, die Adenylatcyclase, die ATP in zyklisches AMP umzuwandelt. 06:08 Dieses zyklische AMP wird dann ein allosterischer Effektor der Proteinkinase A. 06:15 An diesem Punkt wird die Proteinkinase A aktiv und stimuliert die Aktivierung der Fructose-Bisphosphatase 2. 06:22 Dadurch wird das Phosphat aus dem Fructose-2,6-Bisphosphat entfernt. 06:26 Das ist der Prozess, den ich auf der vorherigen Folie beschrieben habe. 06:31 Das Fehlen von Fructose-2.6-Bisphosphat stoppt die Glykolyse und lässt die Gluconeogenese in Gang kommen. 06:40 Die Abläufe, die ich zuvor mit der Bindung von Epinephrin oder Glucagon an den Oberflächenrezeptor der Leberzellen beschrieben habe, finden statt, wenn die Glukosekonzentration in der Blutbahn niedrig ist. 06:50 Wenn die Glukosekonzentration im Blutstrom hoch ist, wird Insulin ausgeschüttet. 06:54 Und Insulin, wie wir in dieser Leberzelle sehen werden kehrt all diese Effekte um. 06:58 Die Bindung von Insulin an den Zelloberflächenrezeptor für Insuline, genannt Insulinrezeptor, bewirkt mehrere Dinge. 07:06 Zunächst einmal wird das G-Protein, das zuvor durch das Adrenalin aktiviert wurde, inaktiviert. 07:11 Das bedeutet, dass die Adenylatcyclase, die zyklisches AMP produziert, nicht aktiviert wird und somit nicht mehr etwas produziert, was die Proteinkinase aktiviert. Die Proteinkinase bleibt daher in einem inaktiven Zustand. 07:26 Wenn die Proteinkinase in einem inaktiven Zustand bleibt, bedeutet das, dass es nichts gibt, was Phosphate auf unser Yin-und-Yang-Enzym überträgt. 07:34 Darüber hinaus stimuliert die Bindung von Insulin an die Insulinrezeptoren die Aktivität eines anderen wichtigen Enzyms, das hier abgebildet ist. Das ist eine insulinstimulierte Phosphatase. 07:46 Und was macht eine Phosphatase? Sie entfernt Phosphate aus Dingen. 07:50 Als Wirkung der insulinstimulierten Phosphatase wird die Fruktose-Biphosphatase 2, die aktiv war, als die Phosphate vorhanden waren, inaktiviert und stattdessen in die andere Form des Yin-und-Yang-Enzyms, die PFK 2, umgewandelt. 08:05 Die PFK 2 nimmt natürlich Fructose-6-Phosphat und fügt ihm ein Phosphat hinzu, um F2.6BP herzustellen. 08:13 F2.6BP aktiviert das Glykolyse-Enzym PFK. Und zur gleichen Zeit hemmt das F2.6BP das Gluconeogenese-Enzym F1.6BPase. 08:25 Infolgedessen beginnt die Glykolyse und die Gluconeogenese wird gestoppt. 08:30 Das macht sehr viel Sinn, denn die Glykolyse beginnt und verbrennt Glukose, weil die Zelle und der Körper versuchen, die Glukosekonzentration zu reduzieren.
About the Lecture
The lecture Hormonal Control by Kevin Ahern, PhD is from the course Metabolic Control.
Included Quiz Questions
Which of the following statements about fructose 2,6-bisphosphate (F2,6BP) is NOT true?
- It is made by PFK1.
- It is broken down by FBPase-2.
- Insulin stimulates its production.
- It favors glycolysis by activating phosphofructokinase (PFK).
Which of the following hormones do NOT play a role in maintaining the blood glucose levels in the body of multicellular organisms?
- Androgen
- Inhibin
- Glucagon
- Insulin
- Epinephrine
Which of the following hormones counteracts the effect of epinephrine?
- Insulin
- Androgen
- Inhibin
- Glucagon
- Hepcidin
Which of the following is true of fructose-2,6-bisphosphate?
- It allosterically affects the activities of phosphofructokinase-1 (PFK-1) and fructose-1,6-bisphosphatase enzymes.
- It allosterically affects the activities of the adenyl cyclase enzyme.
- It allosterically affects the function of insulin hormones.
- It stimulates the production of glucagon and epinephrine hormones.
- It stimulates the production of insulin by the pancreas.
Which of the following statements is NOT true regarding the hormonal regulation of glucose in the body?
- Insulin and epinephrine hormones directly bind to the catalytic sites of the FBPase enzyme and regulate the simultaneous working of gluconeogenesis and glycolysis.
- The enzyme protein kinase A gets activated by the hormones epinephrine and glucagon.
- In response to the high levels of insulin, the G-protein gets inactivated, and then the insulin-stimulated phosphatase enzyme converts FBPase to PFK-2 form and starts the glycolysis pathway.
- The activated FBPase-2 activates gluconeogenesis by synthesis of fructose-6-phosphate from fructose-2,6-bisphosphate.
- The activated protein kinase A enzyme causes the phosphorylation of PFK-2 and activates the FBPase-2 activity.
Which of the following is NOT an endocrine gland?
- Brunner's glands
- Pituitary gland
- Thyroid gland
- Adrenal glands
- Pineal gland
Author of lecture Hormonal Control

Kevin Ahern, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
5 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
