Playlist
Show Playlist
Hide Playlist
Corticopapillary Osmotic Gradient

-
Slides 07 Countercurrent UrinarySystem.pdf
-
Download Lecture Overview
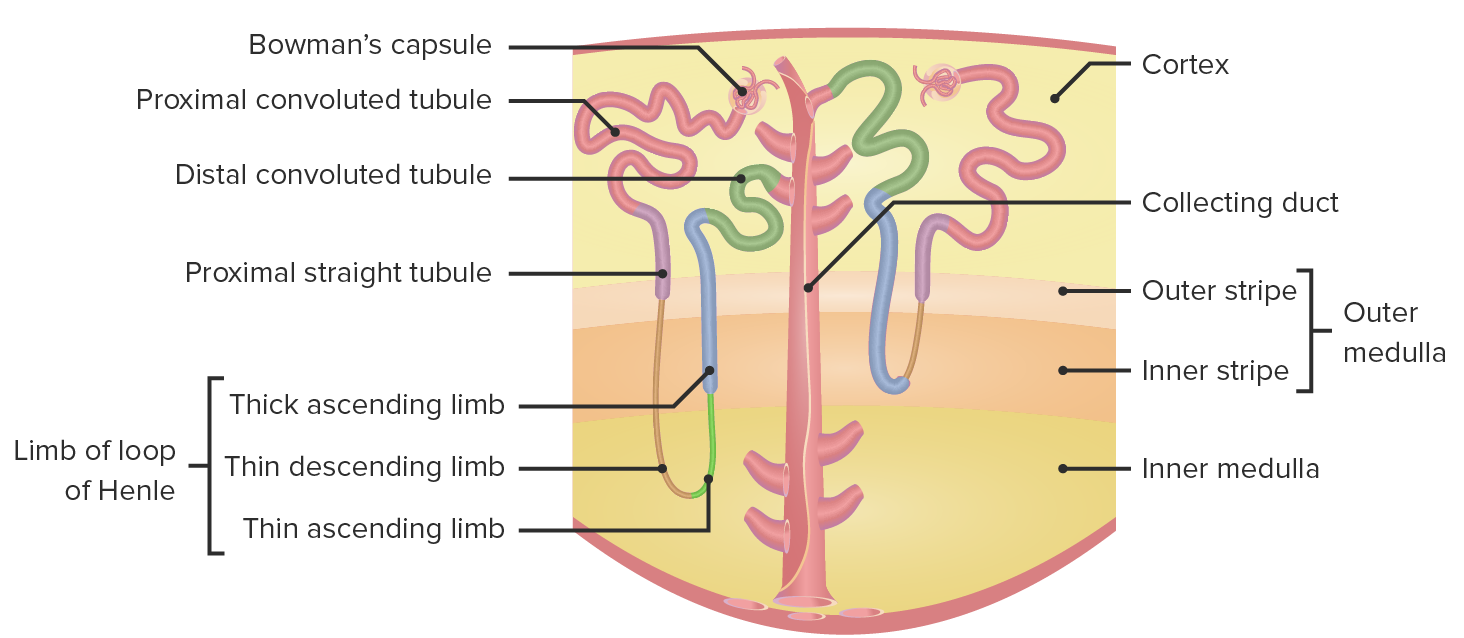
00:01 Now, we’re going to go through a couple of pretty difficult topics, and we’re going to spend a little bit more time trying to unpack them so you can understand them better. 00:10 These are the corticopapillary osmotic gradient in the countercurrent multiplier system. 00:17 To remind you of where we’re talking about in terms of the corticopapillary medullary gradient, as well as the countercurrent multiplier system, this is the loop of Henle. 00:29 In the loop of Henle, we have a thin descending limb, we have the hairpin loop, the thin ascending limb, and the thick ascending limb. 00:40 Notice histologically, these are very different cell types and you notice this because of the thickness of those cells. 00:48 In terms of the thin limbs, they are very narrow in terms of their width, and that is because they don’t do much transport. 00:56 This is a passive process that they’ll be involved with. 00:59 They will either be permeable to a solute, or permeable to water, but not both. 01:04 When we go to the thick ascending limb, you see there’s a number of mitochondria and their nuclei, and you can see all of the different transporters that might be expressed there. 01:14 There will be an active process, meaning that you need to require energy for that transport process to work. 01:21 What is the gradient? We’ve reviewed this earlier, and this involves both an anatomical and osmotic point of view. 01:28 You can see from the cortex all the way to the medullary regions – there is a gradient of osmolality. 01:35 It starts up near 300, which is an iso-osmotic type of condition, all the way down to about 1,200, which is hyper-osmotic. 01:45 Having this gradient present, allows for the various transport processes in the loop of Henle. 01:54 How this is set up? Now, we’re going to use the corticopapillary gradient here on the one side. 02:01 We’ll talk through how the transport process happens along the thin descending limb. 02:07 The thin descending limb is permeable to water. 02:10 So as you travel down towards the hairpin loop turn, more and more water will exit the thin descending limb, and that occurs because there is a gradient. 02:24 That gradient will pull water out to try to equalize. 02:29 So when you’re up on an iso-osmotic condition, there is less water movement. 02:34 When you’re in a hyper-osmotic conditions, there’s a lot of water movement. 02:38 This is all passive, meaning that you don’t require energy for this process to work. 02:45 Then as you travel up back from a hyperosmotic condition to an iso-osmotic condition, we now have the thin ascending limb. 02:57 The think ascending limb is impermeable to water, but it is permeable to solutes such as sodium. 03:05 So if there’s hyper-osmotic conditions, there will be little to no sodium that will leave the thin ascending limb. 03:13 As you go up to more iso-osmotic conditions, that’s when the sodium will leave. 03:18 And this is done in the opposite as the thin descending limb where you have more water traveling out towards the base of the loop. 03:28 In here, you have the solutes traveling out as you start to get more iso-osmotic. 03:35 This is again a passive process losing both water and salt. 03:40 You may think to yourself, “Well, this seems kind of odd. 03:44 Why in the world would you want to lose the water and salt?” You’re losing it out of the tubule lumen, and that means you’re going to be able to reabsorb it. 03:53 So you’re actually protecting the loss of sodium and the loss of water by going through this corticopapillary osmotic gradient. 04:03 Once you reach the thick ascending limb, now you’re going to have an active process to occur. 04:10 What I mean by active is you’ll need sodium and potassium will need to travel via an active sodium-potassium ATPase. 04:19 You’re going to utilize energy for this process to occur. 04:23 The sodium-potassium ATPase located on the basolateral membrane sets up the driving force for transport. 04:30 The major transporters in this area are the triple transporter, which is known as an NCCK transporter. 04:39 It transports sodium, potassium, and 2 chlorides. 04:43 So it involves a lot of co-transport, just with 1 transporter. 04:49 In pharmacology, this is what is going to be the loop sensitive diuretic. 04:55 Now, why are we thinking about this process? We want to reabsorb sodium along with chloride. 05:02 The sodium is then kicked out of the basolateral side of the membrane from the sodium-potassium ATPase – the chloride leaves via a channel. 05:12 What happens to the potassium? It’s complex. It’s goes off both sides. 05:17 So potassium is allowed to move back and forth out of the basolateral membrane, gets pumped back in, then leaves via these ROMK channels, and then travels back in via the triple transporter. 05:29 So potassium is really facilitating the process of reabsorbing sodium and chloride. 05:38 This kind of process, in which you have a triple transporter and you have a sodium-potassium ATPase located on the basolateral membrane, sets up a large electrical gradient from the apical to basolateral side of the membrane. 05:54 The transelectrical gradient is about 7 millivolts. 06:00 This will allow for the movement of ions from the apical to the basolateral side simply by this gradient being present. 06:13 Water doesn’t travel through this particular part; just the ions will travel through. 06:21 This is how both sodium, as well as some potassium, are also reabsorbed. 06:27 If you remember, this is also – if you have other positively charged molecules such as calcium and magnesium, these divalent cations, they can also travel through based upon this gradient, moving through in this paracellular manner, not requiring energy, simply using the gradient to drive the movement. 06:50 So calcium and magnesium, especially magnesium, are reabsorbed by this manner. 06:56 Now, you have the reabsorption of all these ions. 07:01 We talked about this – the peritubular capillaries being very important for this process, the glomerular capillaries being important for the filtration – but what happens towards the loop of Henle? We have a different vessel system that we haven’t discussed very much earlier. 07:19 These are the vasa recta vessels. 07:22 So where do these come from? You have the afferent arteriole delivering blood flow into the glomerular capillary, then you have the efferent arteriole leaving the glomerular capillary going into the peritubular capillary system. 07:38 After the peritubular capillaries, then you have these vasa recta vessels that are traveling in close proximity to the loop of Henle, and this loop is the thin descending and the thin ascending loops. 07:52 These can be seen if we kind of do a small cross-section of these vessels. 07:57 So you have the cortical collecting tubules, the nephron loops, and these vasa recta blood vessels all located in the same area. 08:06 This is why you can utilize a cortical to medullary capillary osmotic gradient so well because you have these vessels traveling in close proximity to the both nephron loops and the collecting tubules. 08:21 This allows for equilibrium to occur very rapidly because of this close proximity.
About the Lecture
The lecture Corticopapillary Osmotic Gradient by Thad Wilson, PhD is from the course Urinary Tract Physiology.
Included Quiz Questions
Which transporter, charged with sodium reabsorption, is on the apical membrane of the thick ascending limb?
- Sodium-chloride-potassium cotransporter
- Sodium-chloride exchanger
- Epithelial sodium channel
- Sodium-potassium ATPase
Which blood vessels are intertwined between the nephron loop and the collecting duct?
- Vasa recta vessels
- Peritubular capillaries
- Glomerular capillaries
- Medullary capillaries
At what corticopapillary osmolality will more water molecules leave the thin descending limb due to passive transport?
- 750
- 300
- 450
- 600
- 500
Which of the following type of transporters is sensitive to loop diuretics?
- NKCC transporter
- Sodium-potassium ATPase
- ROMK
- Chloride channels
- Potassium channels
Which ions can be transported through both in and out of the cell through basolateral and apical membranes in the thick ascending membrane?
- Potassium
- Sodium
- Chloride
- Calcium
- Magnesium
Author of lecture Corticopapillary Osmotic Gradient

Thad Wilson, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
5 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
