Playlist
Show Playlist
Hide Playlist
Class Switch Recombination, Co-Expression and B-Cell Development – Lymphocyte Development

-
Slides Adaptive Immune System.pdf
-
Download Lecture Overview
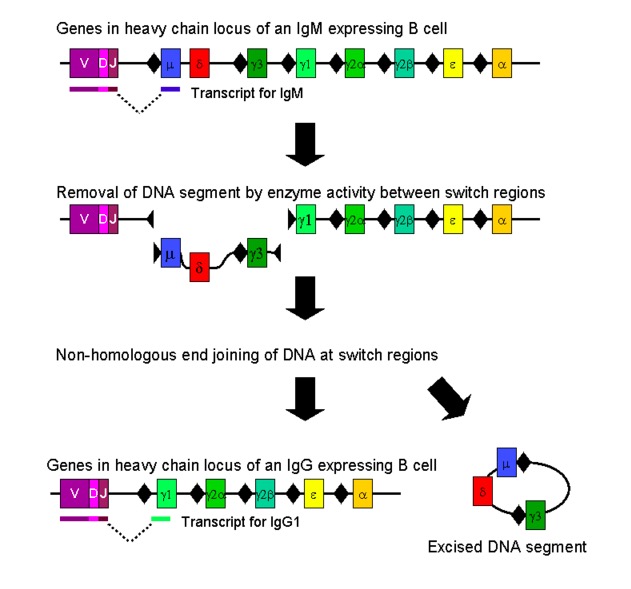
00:01 B-cells initially start off making the IgM class of antibody. 00:05 But following antigen stimulation, they will switch to making other classes of antibody - IgG, IgA or IgE. 00:14 This class switching also takes place at the genetic level. 00:18 Here we can see the immunoglobulin heavy chain gene locus with the VDJ segment already recombined. 00:28 This is lying immediately upstream of C mu (Cµ) and C delta (Cδ). 00:33 So the IgM class of antibody will be produced. 00:38 However, following antigen stimulation, the antibody going to be produced will be, in this instance, an IgE antibody. 00:48 This requires signals from helper T-cells, for example CD40 ligand on the surface of the T-cell interacting with CD40 on the surface of the B-cell. 00:59 And also cytokines produced by the helper T-cells. 01:03 What will happen in this particular case is that the switch sequence immediately upstream of the Cµ gene is going to be placed next to the switch sequence immediately upstream of the C epsilon (Cε) gene. 01:19 So there’s a recombination of the switch sequence for mu with the switch sequence for epsilon, and the deletion of the intervening constant region genes. 01:30 Now there will be transcription and RNA splicing, and the messenger RNA encoding the IgE antibody, in other words, VDJ plus Cε will be translated into protein. 01:46 And plasma cells will secret the IgE class of antibody. 01:52 Naïve B-cells co-express IgM and IgD on their cell surface. 01:58 And when activated, and when they differentiate into plasma cells, those plasma cells also secrete IgM and IgD. 02:06 Although they produce much, much more IgM than they do IgD. 02:10 You’ll hardly be surprised to learn that yet again, it is at the genetic level that this co-expression is determined. 02:17 Here we can see the already recombined VDJ segment in the B-cell immunoglobulin heavy chain DNA. 02:25 And immediately downstream of this recombined VDJ, is the Cµ and Cδ genes. 02:34 It’s very noteworthy that out of the nine Constant region gene segments, all of them contain a switch sequence apart from Cδ. 02:46 And this permits the transcription of both Cµ and Cδ genes together with the VDJ. 02:56 This enables the co-expression of IgM and IgD, which we can see here on the surface of a naïve B-cell. 03:07 The primary RNA transcript containing both the Cµ and Cδ genes is then alternatively spliced to produce messenger RNA encoding Cµ, in other words an IgM antibody; or alternatively spliced to contain Cδ which will encode the IgD antibody. Here we have the typical B-cell receptor comprising a transmembrane version of the antibody molecule. You can see the typical antibody molecule here, with two identical heavy chains, disulphide bonded to each other. And two identical light chains, each of which is disulphide bonded to one of the heavy chains. The antibody molecule sits in the membrane of the B-lymphocyte, held there by a transmembrane sequence. 04:00 Associated with the transmembrane version of the antibody molecule are two molecules called Ig-alpha (Ig-α) and Ig-beta (Ig-β). 04:09 Importantly, in the cytoplasmic domain of these two molecules, there are ITAMs (Immunoreceptor Tyrosine-Based Activation Motifs). 04:21 These are required for the B-cell to signal into the nucleus once antigen is detected. 04:28 This is initiated by Src family kinases, adding phosphates to the tyrosine residues within the ITAMs. 04:45 If we look at the different types of B-cell that exists, there are essentially two fundamentally different types of B-lymphocyte. 04:53 They’re called B1 and B2. 04:57 B1 cells are produced in the liver, particularly in the fetus, and they are self renewing cells, in other words B1 cells will produce more B1 cells. 05:12 They are present in the peritoneal and pleural cavities. 05:16 And they do not require T-cells in order to become activated, in other words they mediate what we call a T-independent response. 05:27 The antibodies they produce are low affinity IgM class antibodies. 05:33 And they’re often referred to as natural antibodies. 05:36 And this term relates to the fact that the antibodies seem to be produced in the absence of any obvious antigenic stimulation. 05:46 However, the majority of B-cells are the B2 cells. 05:51 And in fact, whenever you hear anybody talking about B-cells or B-lymphocytes, these are the cell type that they’re talking about. 05:58 B2 cells rather than B1 cells. 06:01 The B2 cells, as I’ve already mentioned are the conventional B-cells. 06:05 They are replenished from the bone marrow, in other words the bone marrow keeps producing more B-cells as they’re required. 06:13 They are present predominantly within the secondary lymphoid tissues, although you can find them throughout the body. And they require T-cells in order to develop into plasma cells, and particularly for class switching, to produce the IgG, IgA and IgE classes of antibody. 06:32 And they can produce high affinity antibodies of these three classes.
About the Lecture
The lecture Class Switch Recombination, Co-Expression and B-Cell Development – Lymphocyte Development by Peter Delves, PhD is from the course Adaptive Immune System. It contains the following chapters:
- Class Switch Recombination
- Co-Expression on Naive B-Cells
- The B-Cell Receptor
- B1 and B2 Cells
Included Quiz Questions
What are the two classes of antibodies that are expressed on the surface of naive B-cells?
- IgM and IgD
- IgM and IgG
- IgG and IgD
- IgA and IgE
- IgG and IgE
Which of the following initiates/modulates class switching of the B-cell antibodies after antigen exposure?
- Signals from Helper T cells
- Recombination
- Deletion of intervening genes
- RNA splicing
- Translation
How are naive B cells capable of coexpressing IgM and IgD?
- The delta gene immediately downstream from the mu gene does not contain a switch sequence.
- The mu gene immediately upstream from the delta gene does not contain a switch sequence.
- A single gene codes for both IgM and IgD.
- The only genes available on the constant region in naive B cells are the mu and delta genes.
- Gene recombination joins mu and delta genes.
Which of the following is an integral component for the initiation of the signaling pathway and is found in the cytoplasmic tails of cell surface proteins?
- Immunoreceptor tyrosine-based activation motif (ITAM)
- Heavy chain gene sequences
- The transmembrane sequence that holds antibody into the B cell membrane
- Light chain gene sequences
- Recombination signal sequences
Author of lecture Class Switch Recombination, Co-Expression and B-Cell Development – Lymphocyte Development

Peter Delves, PhD
Customer reviews
5,0 of 5 stars
| 5 Stars |
|
1 |
| 4 Stars |
|
0 |
| 3 Stars |
|
0 |
| 2 Stars |
|
0 |
| 1 Star |
|
0 |
Dr Delves is an excellent lecturer. He makes everything simple and understandable. I like the fact that he raises his voice to indicate key words. I have already recommended the whole curriculum to Medical Students in other countries
